

Review
doi: 10.1007/128_2012_412.
Protein dynamics and the enzymatic reaction coordinate
Affiliations
- PMID: 23508766
- PMCID: PMC4346281
- DOI: 10.1007/128_2012_412
Item in Clipboard
Review
Protein dynamics and the enzymatic reaction coordinate
Top Curr Chem.
2013.
Abstract
This chapter discusses progress over the past 15 years in understanding the role of protein dynamics in enzymatically catalyzed chemical reactions. Research has shown that protein motion on all timescales from femtoseconds to milliseconds can contribute to function, and in particular in some enzymes there are sub-picosecond motions, on the same timescale as barrier passage, the couple directly to chemical transformation, and are thus part of the reaction coordinate. Approaches such as transition path sampling and committor analysis have greatly enhanced our understanding of these processes.
Figures
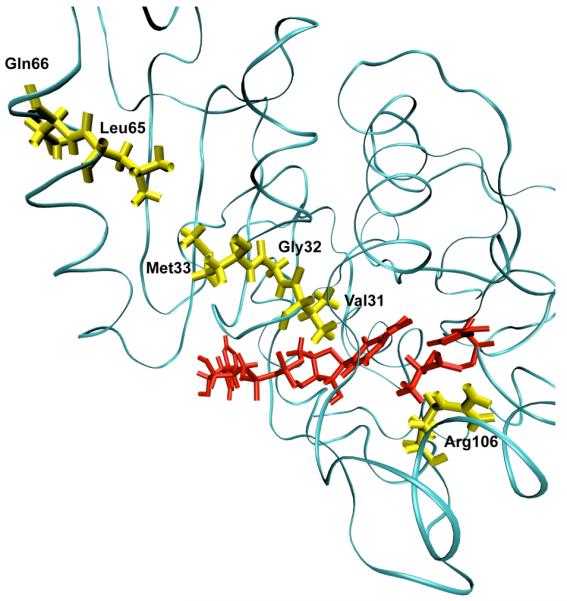
A ribbon diagram of a monomer of human heart lactate dehydrogenase demonstrating the promoting vibration. The donor and acceptor: the nicotinamide ring, and lactate are colored red. The promoting vibration residues, Val31, Gly32, Met33, Leu65, Gln66 (yellow) are compressed towards the active site bringing the NC4 of the nicotinamide ring and substrate carbon closer together while Arg106 (yellow) relaxes away locking the substrate in product formation. These residues span the entire length of the monomer to the edge of the protein. Reprinted with permission from J. Chem. Phys. B. 10.1021/jp207876k 2011
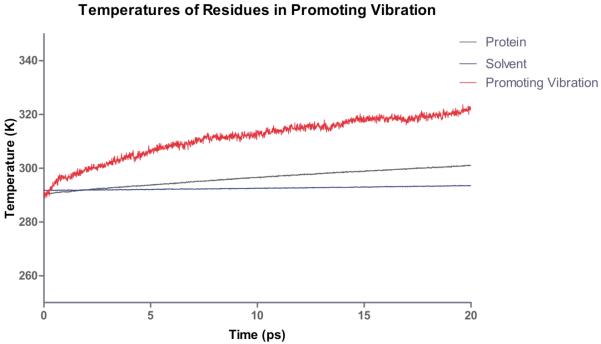
Computational heating experiment of the active site of LDH. The nicotinamide ring is heated and the temperature of the solvent, protein and residues along the promoting vibration are monitored. The promoting vibration is clearly hotter than the rest of the protein indicating a preferred direction of thermal energy transfer. Reprinted with permission from J. Chem. Phys. B. 10.1021/jp207876k 2011
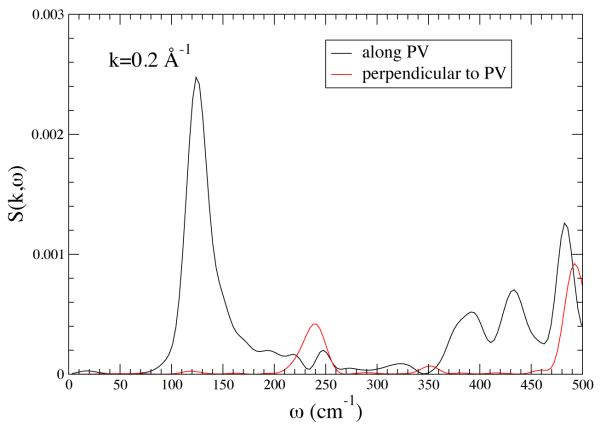
The structure factor S(k,ω) for k = 0.2 A°−1 along the PV axis (black line) and along an axis perpendicular to the promoting vibration. There is strong anisotropy. The sharpness of the peak at about 150 cm−1 means that there are stable fluctuations along the PV axis for that frequency. Reprinted with permission from J. Chem. Phys. B. 10.1021/jp210347h 2011
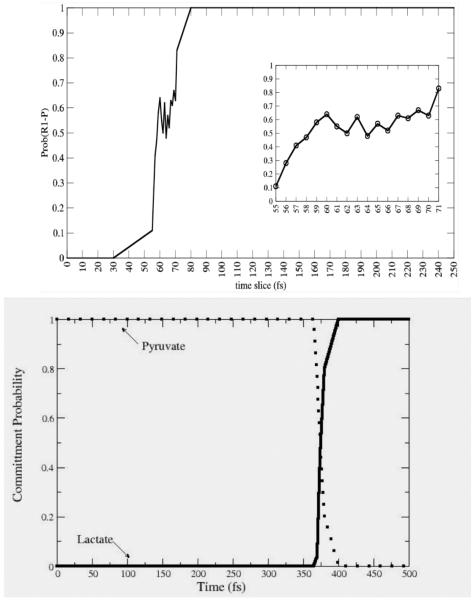
Commitment probability values were calculated for both PNP (top panel) and LDH (lower panel) along sample trajectories from the reactive path ensembles for the respective enzymes. It is clear that the transition region for LDH is far shorter in time duration than for PNP, but even for PNP, the residence time is on the order of 10-20 fsecs. Reprinted with permission from J. Chem. Phys. B. 10.1021/jp207876k 2011
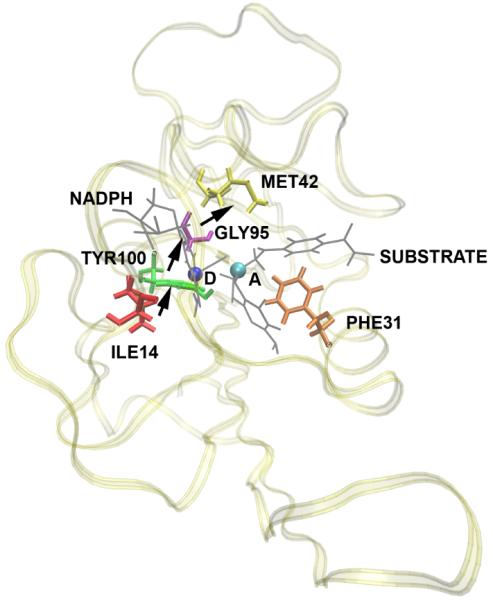
A representation of the direction of motion of residues found in DHFR to be concurrent with the reaction. D is the donor and A is the acceptor. The substrate and the NADPH molecules are shown in gray and the rest of the protein is shown as ribbons. Subsequent committor analysis showed that this motion was not in fact part of the reaction coordinate, and so not a causal part of the reaction mechanism.
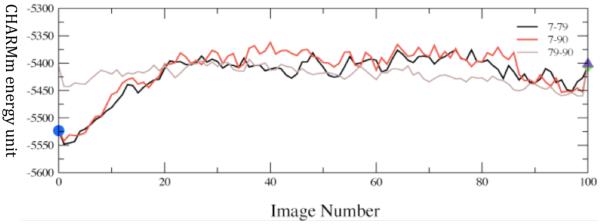
Converged average potential energies between stable, low-distance donor acceptor conformations. The energies are computed using finite temperature string methods. Numerical labels are arbitrary names given to clusters of conformations that structural analysis showed to be identical. The x-axis, which denotes conformational variation, is the string image number. Reprinted with permission from J. Chem. Phys. B. 10.1021/jp207876k 2011
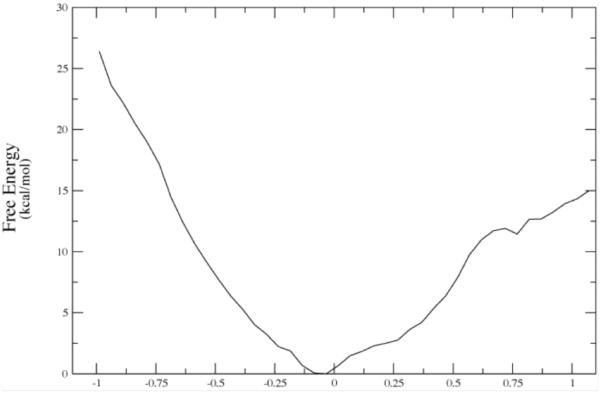
The converged free energy corresponding to one of the potential energy curves above (red curve labeled 7-90.) Note that the minima in average potential energy are on the edges of a rising free energy. The minima on Figure 6a given in string images correspond to −.50 and + .69 angstroms in Δ-RMSD units along the string coordinate on 6b. Reprinted with permission from J. Chem. Phys. B. 10.1021/jp207876k 2011
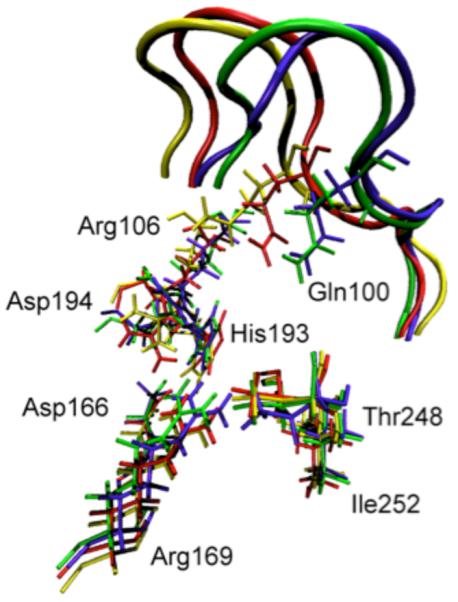
Overlay of the active site residues of 4 representative potential energy stable conformations (three of which are part of Figure 5a.) Residues belonging to the same conformation are shown with the same color. At the top of the picture, the active site loop is shown (residues 96–106, its residues 100 and 106 are shown explicitly) The 4 residues at the bottom of the figure are related to each other by a simple translation. However, the important residues His193, Arg106, Gln100 and Asp194 are being pushed by the active site loop (which has a different closed position in each of the 4 conformations), and their geometric arrangement is different in a non-trivial way among the 4 conformations. Transitions between these conformations were seen to always involve partial loop opening. Reprinted with permission from J. Chem. Phys. B. 10.1021/jp207876k 2011
References
-
- Austin RH, Beeson KW, Eisenstein L, Frauenfe1der H, Gunsalus IC. Biochemistry. 1975;14:5355. - PubMed
-
- Agmon N, Hopfield J. J. Chern. Phys. 1983;79:2042.
-
- Brooks B, Bruccoleri R, Olafson B, States D, Swaminathan S, Karplus M. J. Comp. Chem. 1983;4:187–217.
-
- Gao J, Amara P, Alhambra C, Field M. J. Phys. Chem. A. 1998;102:4714.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
