

Beet yellows virus replicase and replicative compartments: parallels with other RNA viruses
- PMID: 23508802
- PMCID: PMC3589766
- DOI: 10.3389/fmicb.2013.00038
Beet yellows virus replicase and replicative compartments: parallels with other RNA viruses
Abstract
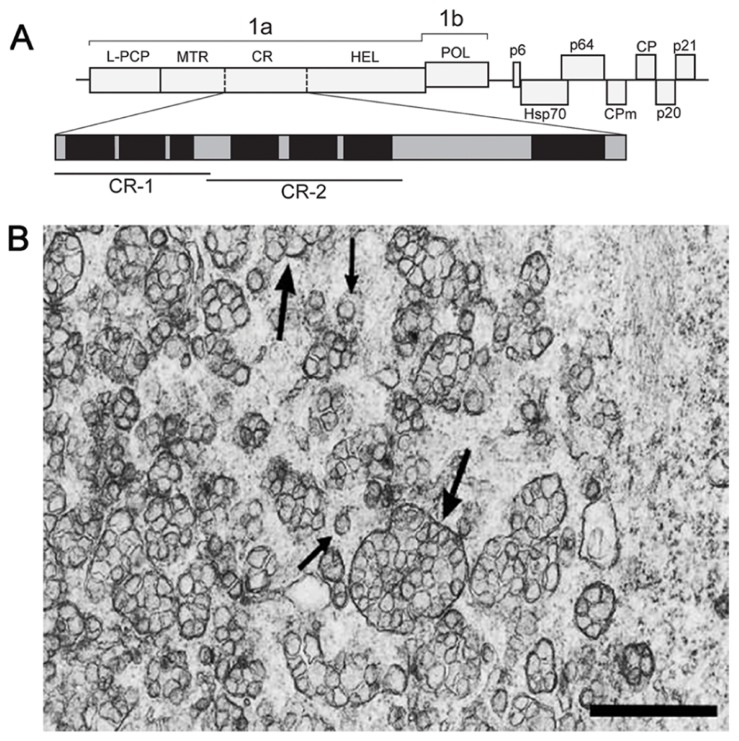
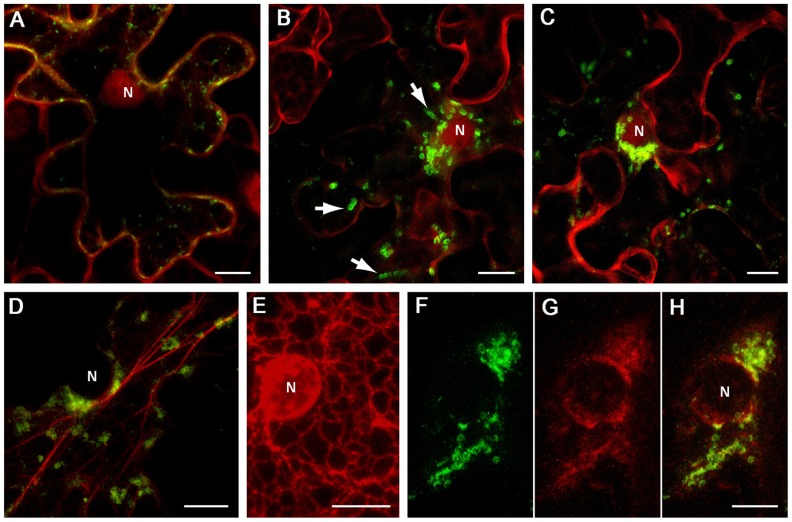
In eukaryotic virus systems, infection leads to induction of membranous compartments in which replication occurs. Virus-encoded subunits of the replication complex mediate its interaction with membranes. As replication platforms, RNA viruses use the cytoplasmic surfaces of different membrane compartments, e.g., endoplasmic reticulum (ER), Golgi, endo/lysosomes, mitochondria, chloroplasts, and peroxisomes. Closterovirus infections are accompanied by formation of multivesicular complexes from cell membranes of ER or mitochondrial origin. So far the mechanisms for vesicles formation have been obscure. In the replication-associated 1a polyprotein of Beet yellows virus (BYV) and other closteroviruses, the region between the methyltransferase and helicase domains (1a central region (CR), 1a CR) is marginally conserved. Computer-assisted analysis predicts several putative membrane-binding domains in the BYV 1a CR. Transient expression of a hydrophobic segment (referred to here as CR-2) of the BYV 1a in Nicotiana benthamiana led to reorganization of the ER and formation of ~1-μm mobile globules. We propose that the CR-2 may be involved in the formation of multivesicular complexes in BYV-infected cells. This provides analogy with membrane-associated proteins mediating the build-up of "virus factories" in cells infected with diverse positive-strand RNA viruses (alpha-like viruses, picorna-like viruses, flaviviruses, and nidoviruses) and negative-strand RNA viruses (bunyaviruses).
Keywords: RNA virus replication; endoplasmic reticulum modification; intracellular traffic; membrane vesicles; virus replication factory.
Figures
Similar articles
-
Inspirations on Virus Replication and Cell-to-Cell Movement from Studies Examining the Cytopathology Induced by Lettuce infectious yellows virus in Plant Cells.Front Plant Sci. 2017 Sep 27;8:1672. doi: 10.3389/fpls.2017.01672. eCollection 2017. Front Plant Sci. 2017. PMID: 29021801 Free PMC article. Review.
-
A conserved region in the Closterovirus 1a polyprotein drives extensive remodeling of endoplasmic reticulum membranes and induces motile globules in Nicotiana benthamiana cells.Virology. 2017 Feb;502:106-113. doi: 10.1016/j.virol.2016.12.006. Epub 2016 Dec 25. Virology. 2017. PMID: 28027478
-
Beet yellows closterovirus: complete genome structure and identification of a leader papain-like thiol protease.Virology. 1994 Jan;198(1):311-24. doi: 10.1006/viro.1994.1034. Virology. 1994. PMID: 8259666
-
Complete sequence of the citrus tristeza virus RNA genome.Virology. 1995 Apr 20;208(2):511-20. doi: 10.1006/viro.1995.1182. Virology. 1995. PMID: 7747424
-
Viral RNA replication in association with cellular membranes.Curr Top Microbiol Immunol. 2005;285:139-73. doi: 10.1007/3-540-26764-6_5. Curr Top Microbiol Immunol. 2005. PMID: 15609503 Free PMC article. Review.
Cited by
-
Double-Membrane Vesicles as Platforms for Viral Replication.Trends Microbiol. 2020 Dec;28(12):1022-1033. doi: 10.1016/j.tim.2020.05.009. Epub 2020 Jun 11. Trends Microbiol. 2020. PMID: 32536523 Free PMC article. Review.
-
DENV up-regulates the HMG-CoA reductase activity through the impairment of AMPK phosphorylation: A potential antiviral target.PLoS Pathog. 2017 Apr 6;13(4):e1006257. doi: 10.1371/journal.ppat.1006257. eCollection 2017 Apr. PLoS Pathog. 2017. PMID: 28384260 Free PMC article.
-
Inspirations on Virus Replication and Cell-to-Cell Movement from Studies Examining the Cytopathology Induced by Lettuce infectious yellows virus in Plant Cells.Front Plant Sci. 2017 Sep 27;8:1672. doi: 10.3389/fpls.2017.01672. eCollection 2017. Front Plant Sci. 2017. PMID: 29021801 Free PMC article. Review.
-
Walking Together: Cross-Protection, Genome Conservation, and the Replication Machinery of Citrus tristeza virus.Viruses. 2020 Nov 26;12(12):1353. doi: 10.3390/v12121353. Viruses. 2020. PMID: 33256049 Free PMC article. Review.
-
Structure and Expression of Large (+)RNA Genomes of Viruses of Higher Eukaryotes.Biochemistry (Mosc). 2021 Mar;86(3):248-261. doi: 10.1134/S0006297921030020. Biochemistry (Mosc). 2021. PMID: 33838627 Free PMC article. Review.
References
-
- Agranovsky A. A., Koonin E. V., Boyko V. P., Maiss E., Froetschl R., Lunina N. A., et al. (1994). Beet yellows closterovirus: complete genome structure and identification of a leader papain-like thiol protease. Virology 198 311–324 - PubMed
-
- Bamunusinghe D., Hemenway C. L., Nelson R. S., Sanderfoot A. A., Ye C. M., Silva M. A., et al. (2009). Analysis of potato virus X replicase and TGBp3 subcellular locations. Virology 393 272–285 - PubMed
-
- Bartenschlager R., Miller S. (2008). Molecular aspects of Dengue virus replication. Future Microbiol. 3 155–165 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials