

Activity-enhancing mutations in an E3 ubiquitin ligase identified by high-throughput mutagenesis
- PMID: 23509263
- PMCID: PMC3619334
- DOI: 10.1073/pnas.1303309110
Activity-enhancing mutations in an E3 ubiquitin ligase identified by high-throughput mutagenesis
Abstract
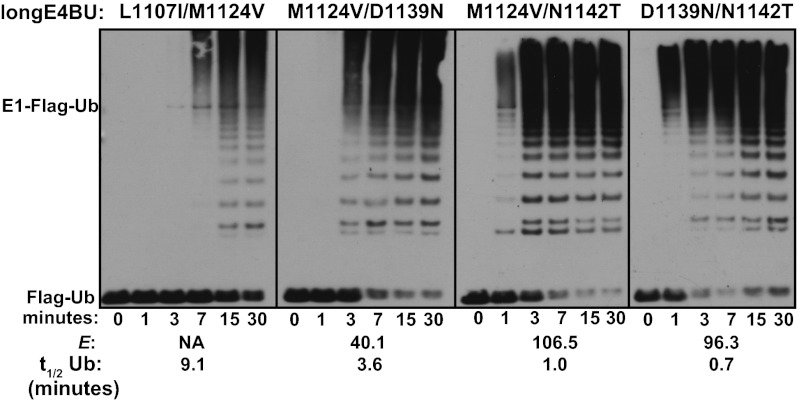
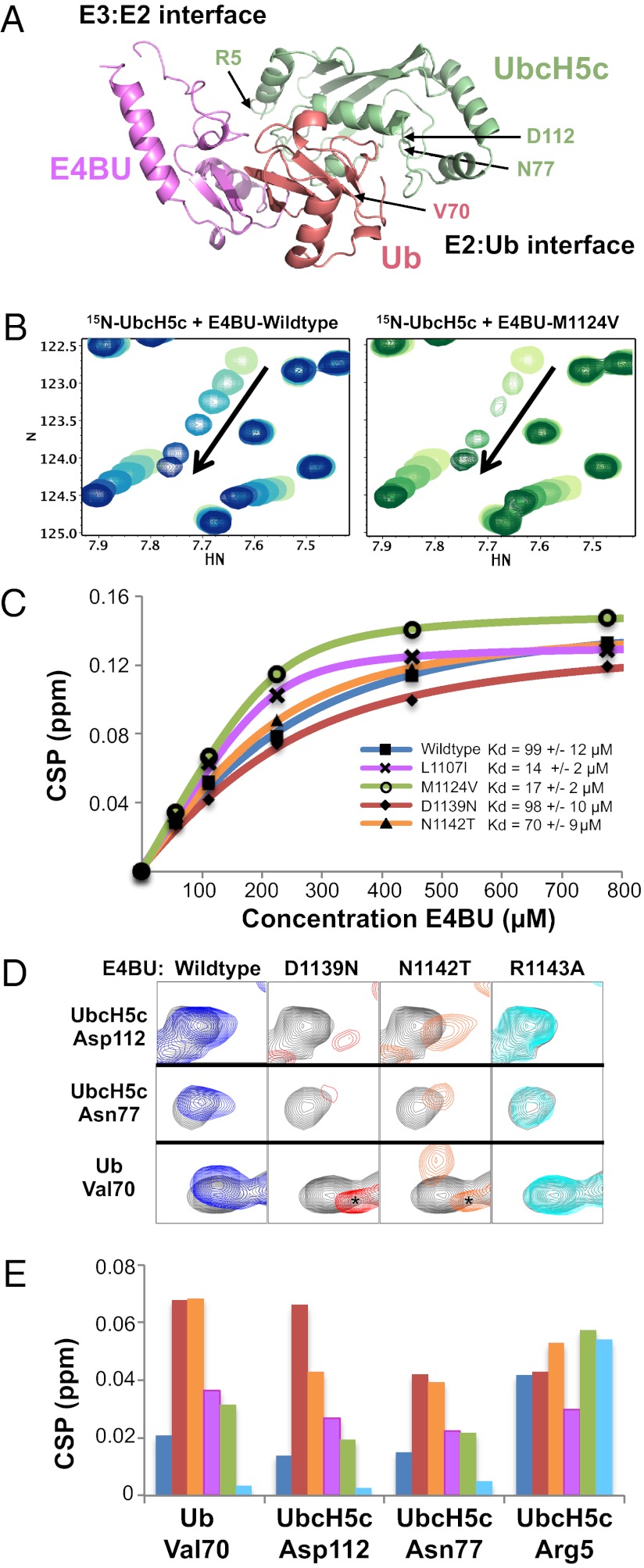
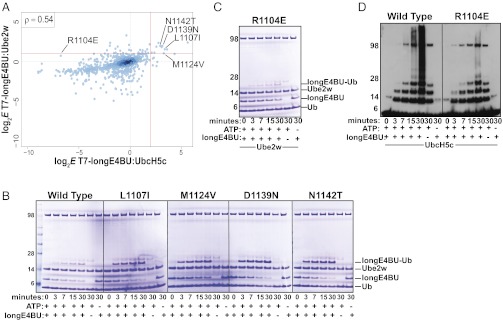
Although ubiquitination plays a critical role in virtually all cellular processes, mechanistic details of ubiquitin (Ub) transfer are still being defined. To identify the molecular determinants within E3 ligases that modulate activity, we scored each member of a library of nearly 100,000 protein variants of the murine ubiquitination factor E4B (Ube4b) U-box domain for auto-ubiquitination activity in the presence of the E2 UbcH5c. This assay identified mutations that enhance activity both in vitro and in cellular p53 degradation assays. The activity-enhancing mutations fall into two distinct mechanistic classes: One increases the U-box:E2-binding affinity, and the other allosterically stimulates the formation of catalytically active conformations of the E2∼Ub conjugate. The same mutations enhance E3 activity in the presence of another E2, Ube2w, implying a common allosteric mechanism, and therefore the general applicability of our observations to other E3s. A comparison of the E3 activity with the two different E2s identified an additional variant that exhibits E3:E2 specificity. Our results highlight the general utility of high-throughput mutagenesis in delineating the molecular basis of enzyme activity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
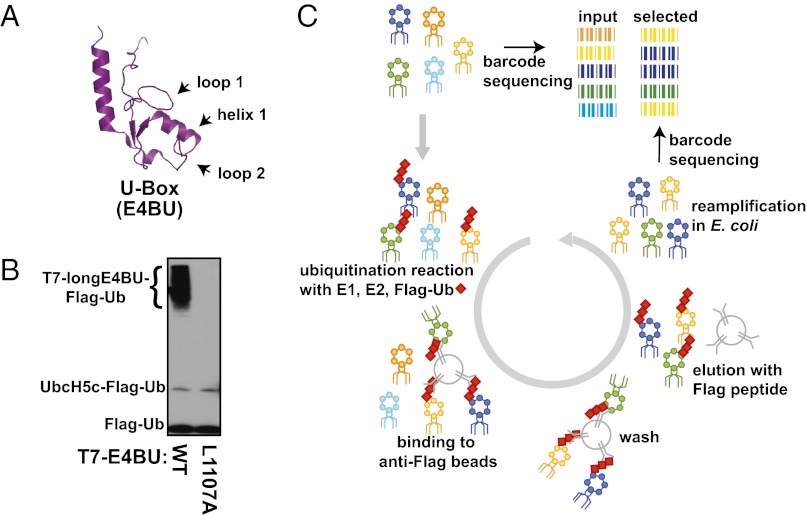
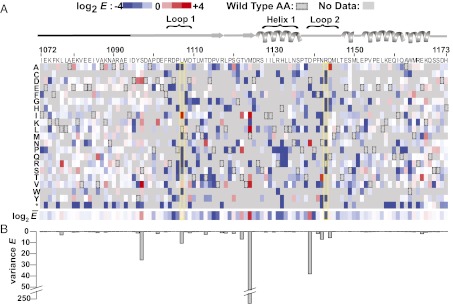
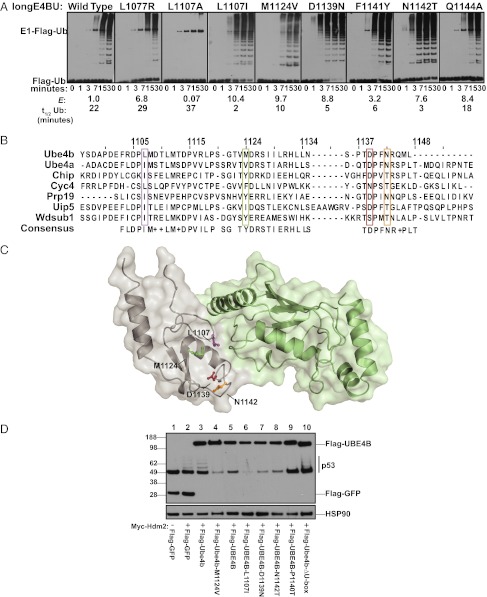
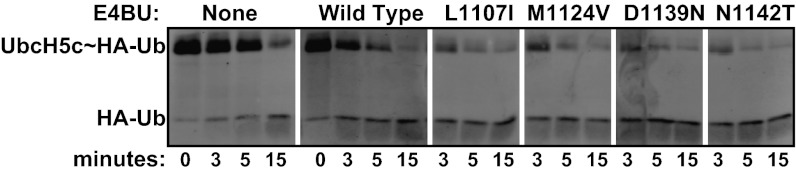
References
-
- Pickart CM, Eddins MJ. Ubiquitin: Structures, functions, mechanisms. Biochim Biophys Acta. 2004;1695(1-3):55–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
