

Conformational plasticity of the type I maltose ABC importer
- PMID: 23509285
- PMCID: PMC3619289
- DOI: 10.1073/pnas.1217745110
Conformational plasticity of the type I maltose ABC importer
Abstract
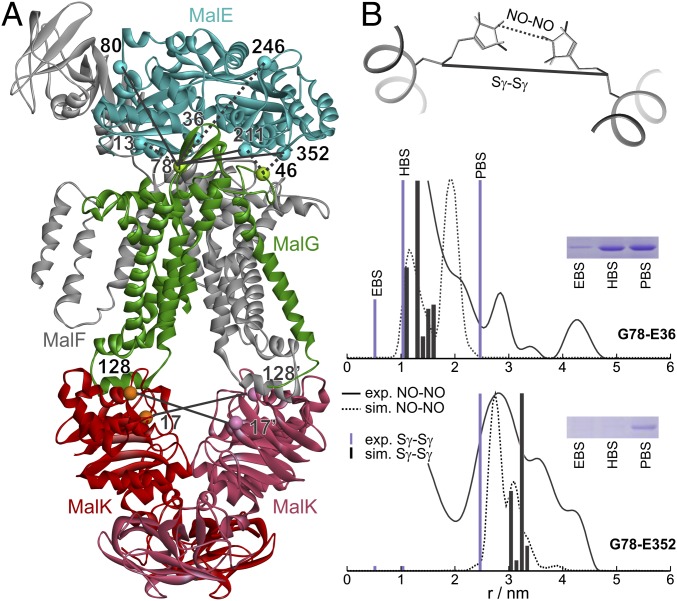
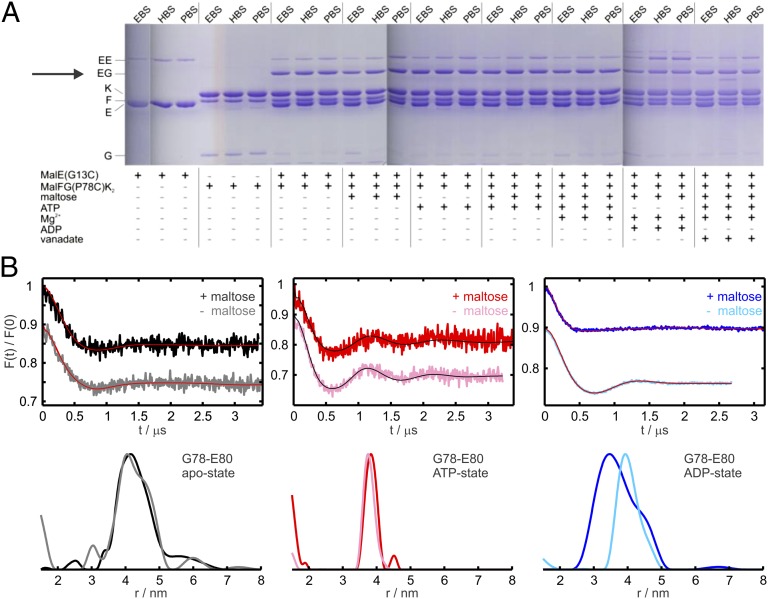
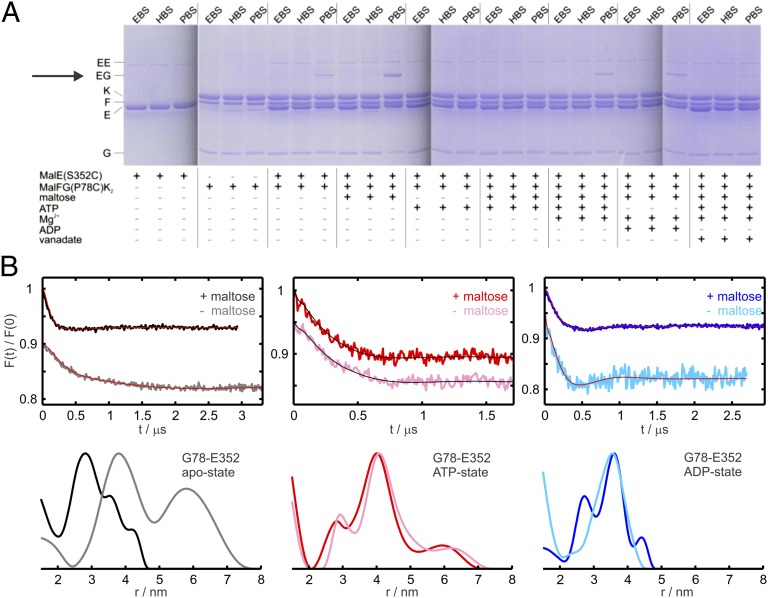
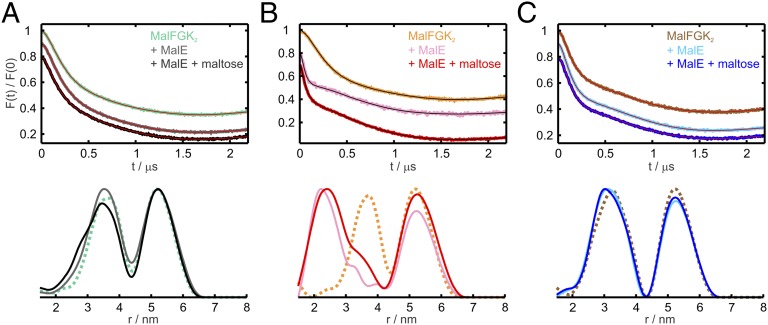
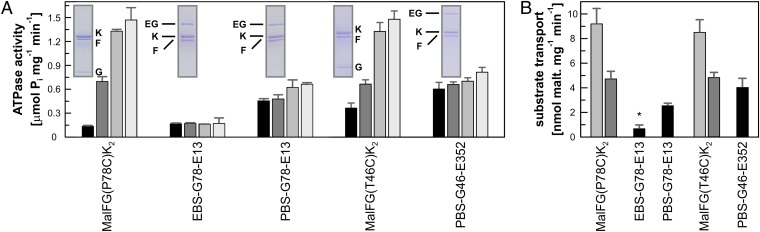
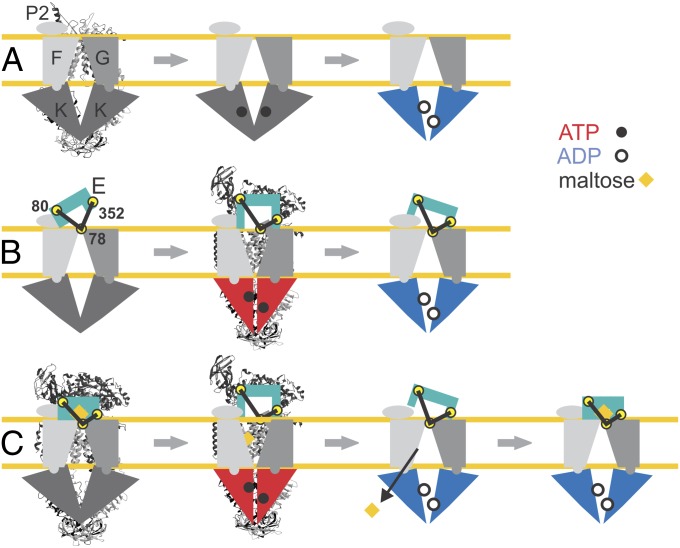
ATP-binding cassette (ABC) transporters couple the translocation of solutes across membranes to ATP hydrolysis. Crystal structures of the Escherichia coli maltose importer (MalFGK2) in complex with its substrate binding protein (MalE) provided unprecedented insights in the mechanism of substrate translocation, leaving the MalE-transporter interactions still poorly understood. Using pulsed EPR and cross-linking methods we investigated the effects of maltose and MalE on complex formation and correlated motions of the MalK2 nucleotide-binding domains (NBDs). We found that both substrate-free (open) and liganded (closed) MalE interact with the transporter with similar affinity in all nucleotide states. In the apo-state, binding of open MalE occurs via the N-lobe, leaving the C-lobe disordered, but upon maltose binding, closed MalE associates tighter to the transporter. In both cases the NBDs remain open. In the presence of ATP, the transporter binds both substrate-free and liganded MalE, both inducing the outward-facing conformation trapped in the crystal with open MalE at the periplasmic side and NBDs tightly closed. In contrast to ATP, ADP-Mg(2+) alone is sufficient to induce a semiopen conformation in the NBDs. In this nucleotide-driven state, the transporter binds both open and closed MalE with slightly different periplasmic configurations. We also found that dissociation of MalE is not a required step for substrate translocation since a supercomplex with MalE cross-linked to MalG retains the ability to hydrolyze ATP and to transport maltose. These features of MalE-MalFGK2 interactions highlight the conformational plasticity of the maltose importer, providing insights into the ATPase stimulation by unliganded MalE.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Moussatova A, Kandt C, O’Mara ML, Tieleman DP. ATP-binding cassette transporters in Escherichia coli. Biochim Biophys Acta. 2008;1778(9):1757–1771. - PubMed
-
- Eitinger T, Rodionov DA, Grote M, Schneider E. Canonical and ECF-type ATP-binding cassette importers in prokaryotes: Diversity in modular organization and cellular functions. FEMS Microbiol Rev. 2011;35(1):3–67. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
