

Network balance via CRY signalling controls the Arabidopsis circadian clock over ambient temperatures
- PMID: 23511208
- PMCID: PMC3619941
- DOI: 10.1038/msb.2013.7
Network balance via CRY signalling controls the Arabidopsis circadian clock over ambient temperatures
Abstract
Circadian clocks exhibit 'temperature compensation', meaning that they show only small changes in period over a broad temperature range. Several clock genes have been implicated in the temperature-dependent control of period in Arabidopsis. We show that blue light is essential for this, suggesting that the effects of light and temperature interact or converge upon common targets in the circadian clock. Our data demonstrate that two cryptochrome photoreceptors differentially control circadian period and sustain rhythmicity across the physiological temperature range. In order to test the hypothesis that the targets of light regulation are sufficient to mediate temperature compensation, we constructed a temperature-compensated clock model by adding passive temperature effects into only the light-sensitive processes in the model. Remarkably, this model was not only capable of full temperature compensation and consistent with mRNA profiles across a temperature range, but also predicted the temperature-dependent change in the level of LATE ELONGATED HYPOCOTYL, a key clock protein. Our analysis provides a systems-level understanding of period control in the plant circadian oscillator.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
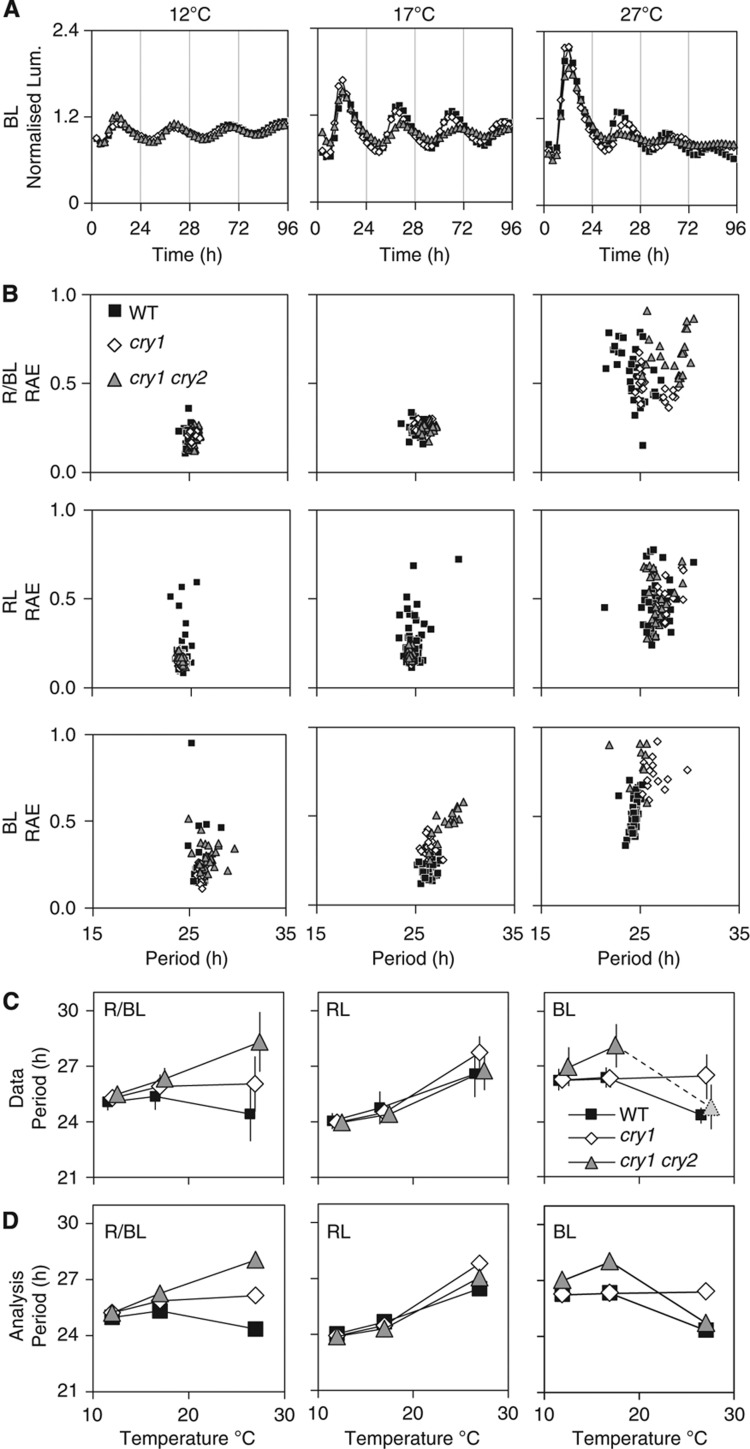
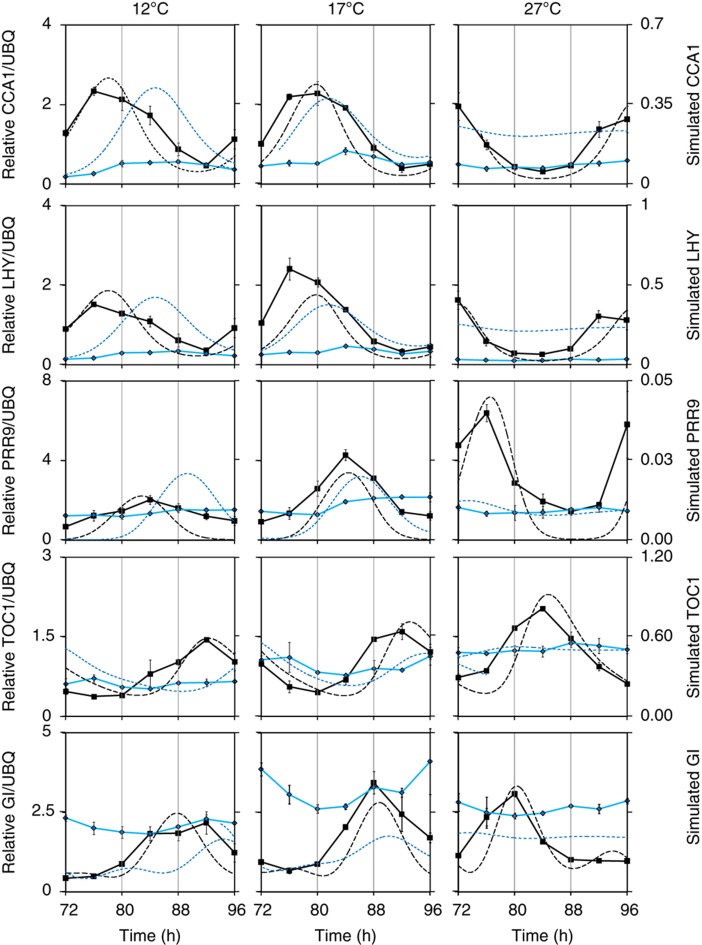
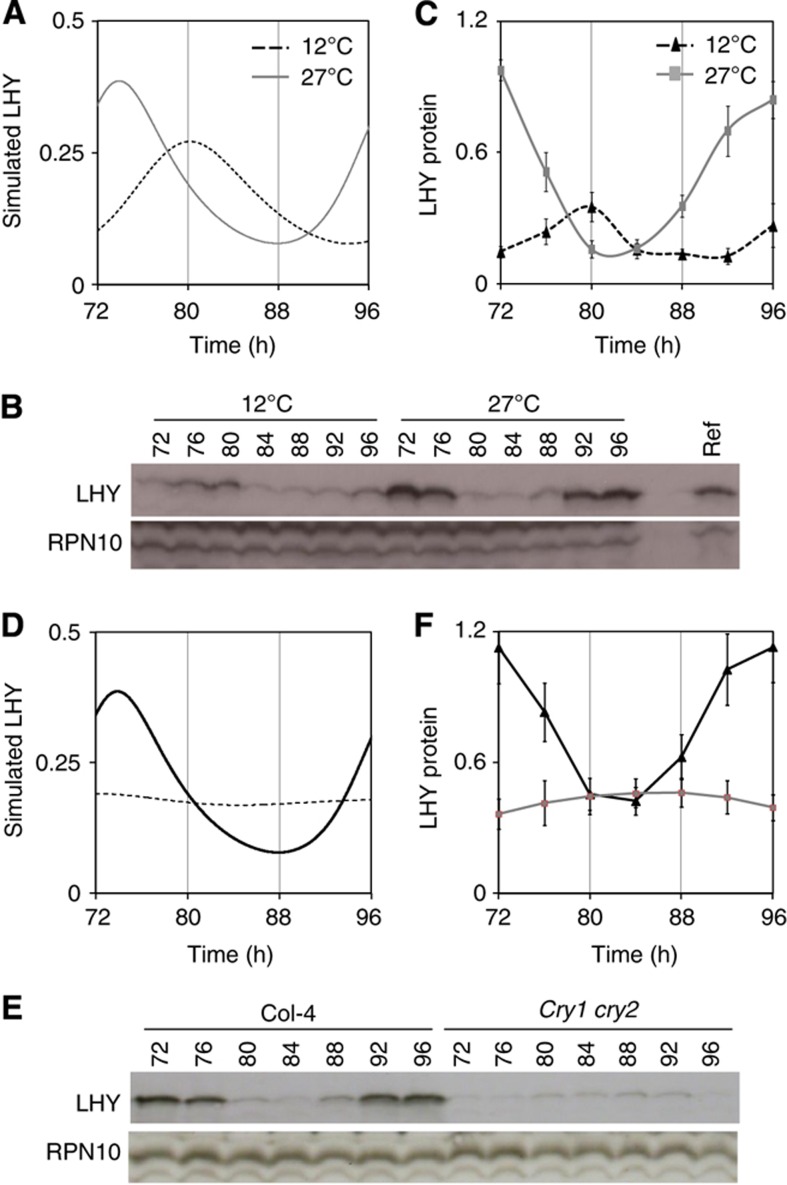
References
-
- Anderson C, Wilkins M (1989) Control of the circadian-rhythm of carbon-dioxide assimilation in Bryophyllum leaves by exposure to darkness and high-carbon dioxide concentrations. Planta 177: 401–408 - PubMed
-
- Bates DM, Maechler M (2011) lme4: Linear Mixed-effects Models Using S4 Classes. R package version 0 999375–37/r1160
Publication types
MeSH terms
Substances
Grants and funding
- BB/D019621/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F005237/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F005318/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F005296/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F005261/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
