

Mechanism of an ATP-independent protein disaggregase: II. distinct molecular interactions drive multiple steps during aggregate disassembly
- PMID: 23519468
- PMCID: PMC3650381
- DOI: 10.1074/jbc.M113.462861
Mechanism of an ATP-independent protein disaggregase: II. distinct molecular interactions drive multiple steps during aggregate disassembly
Abstract
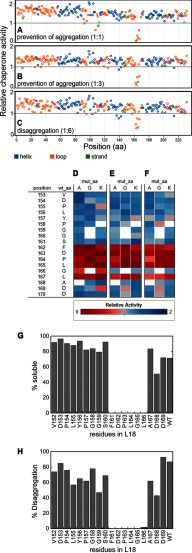
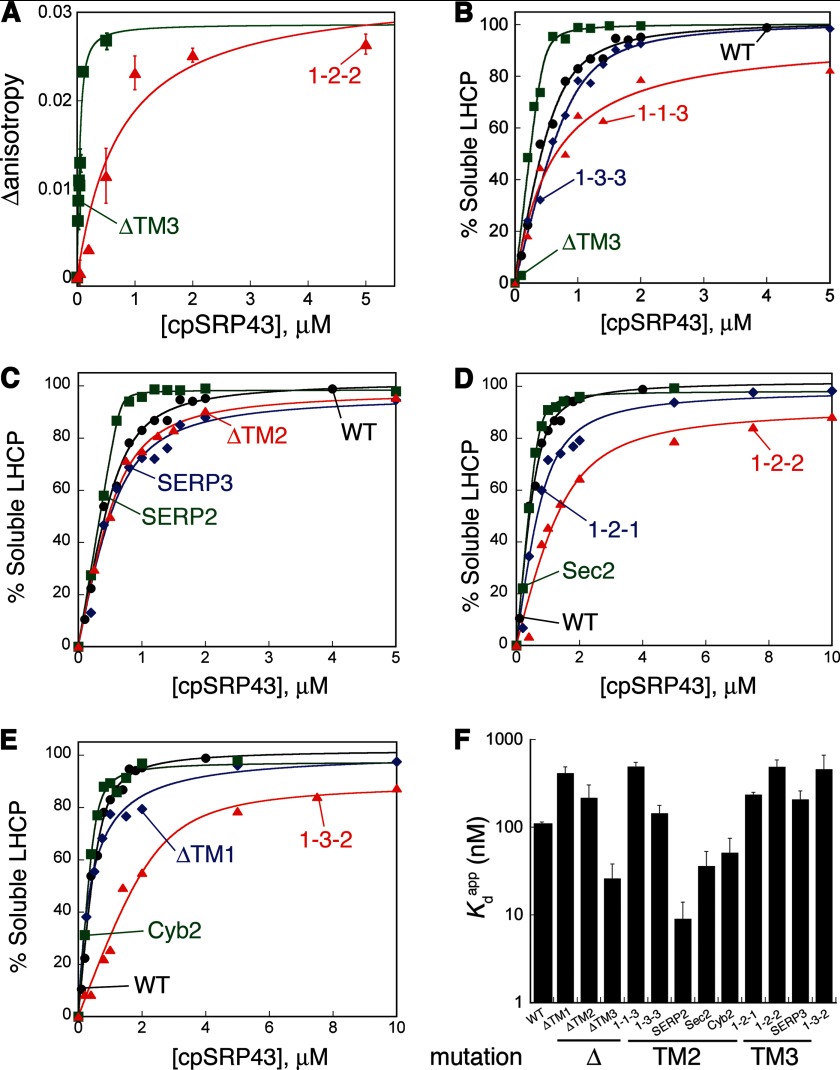
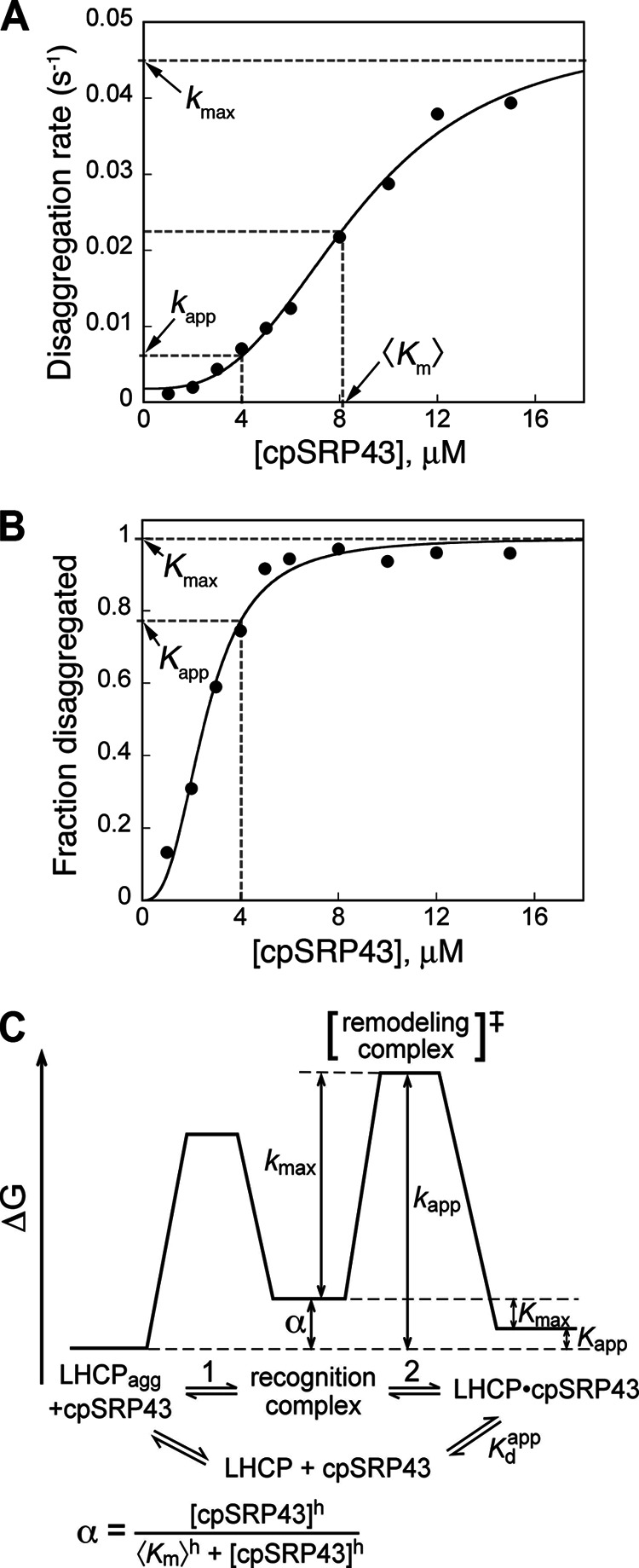
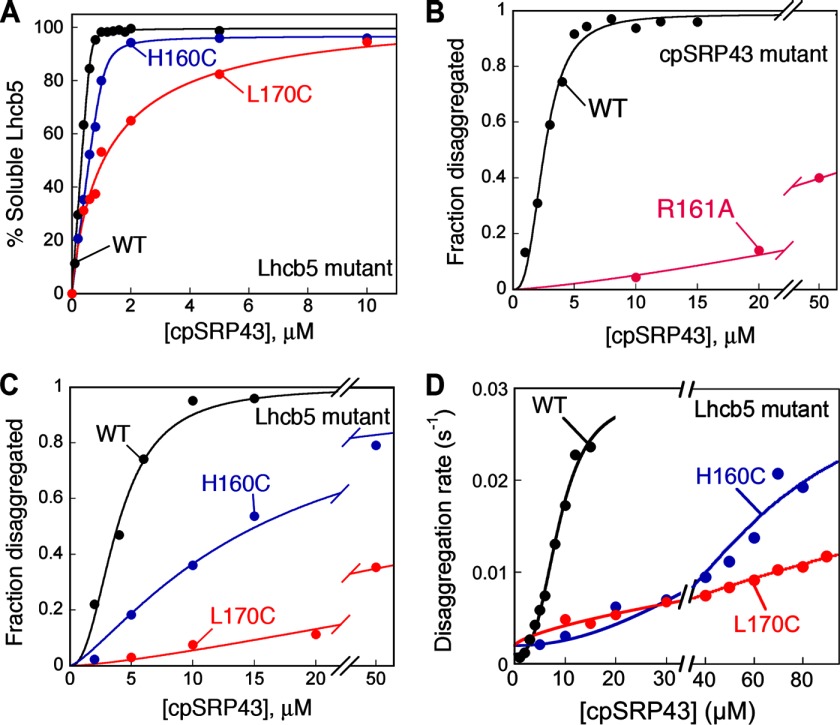
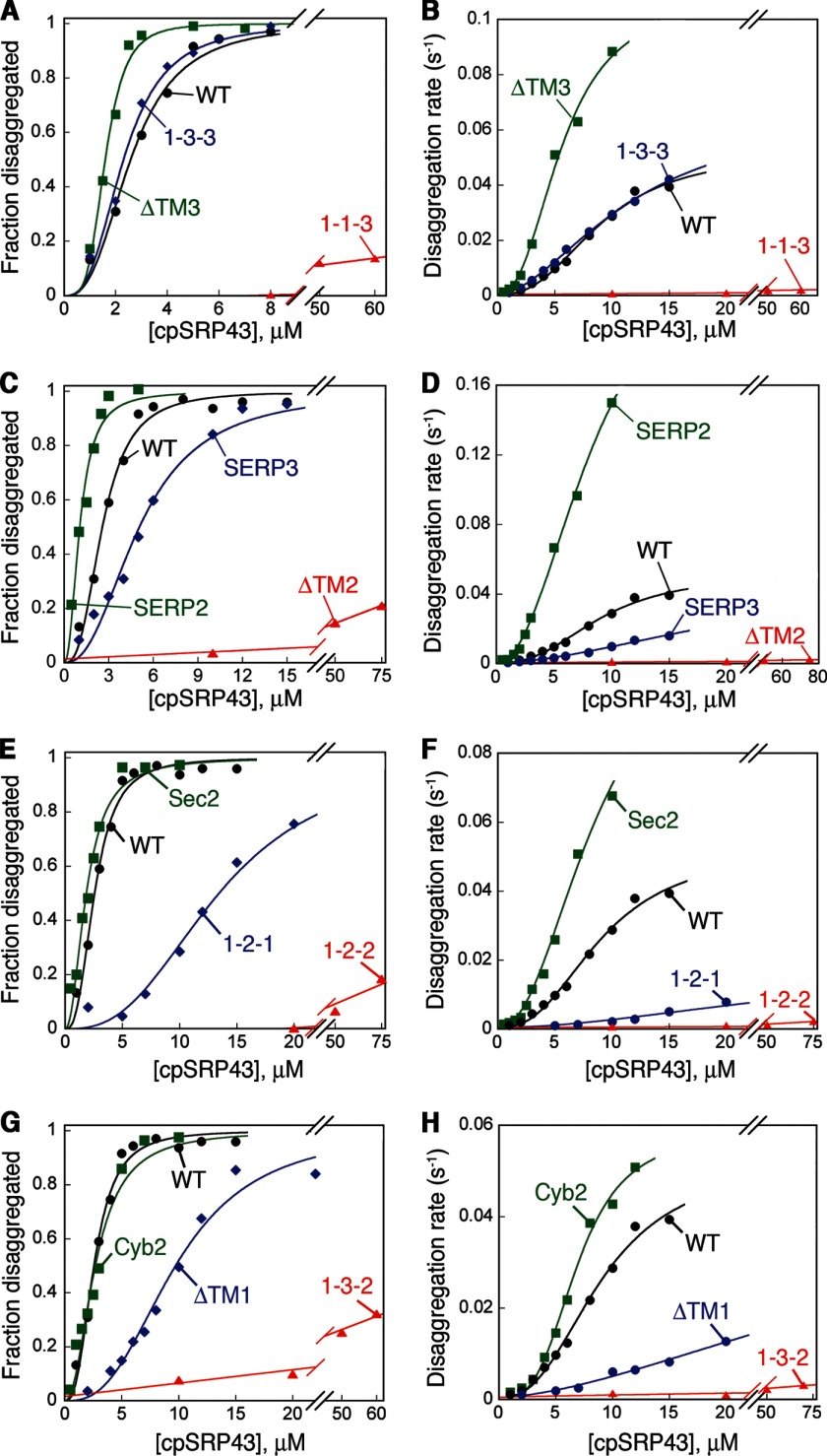
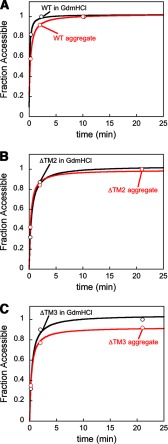
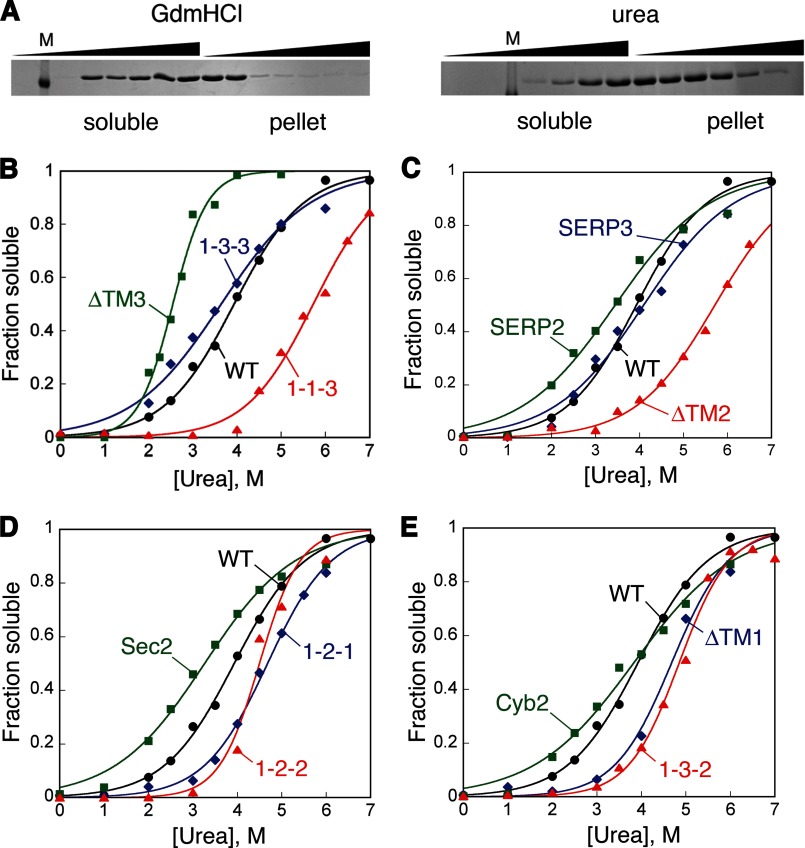
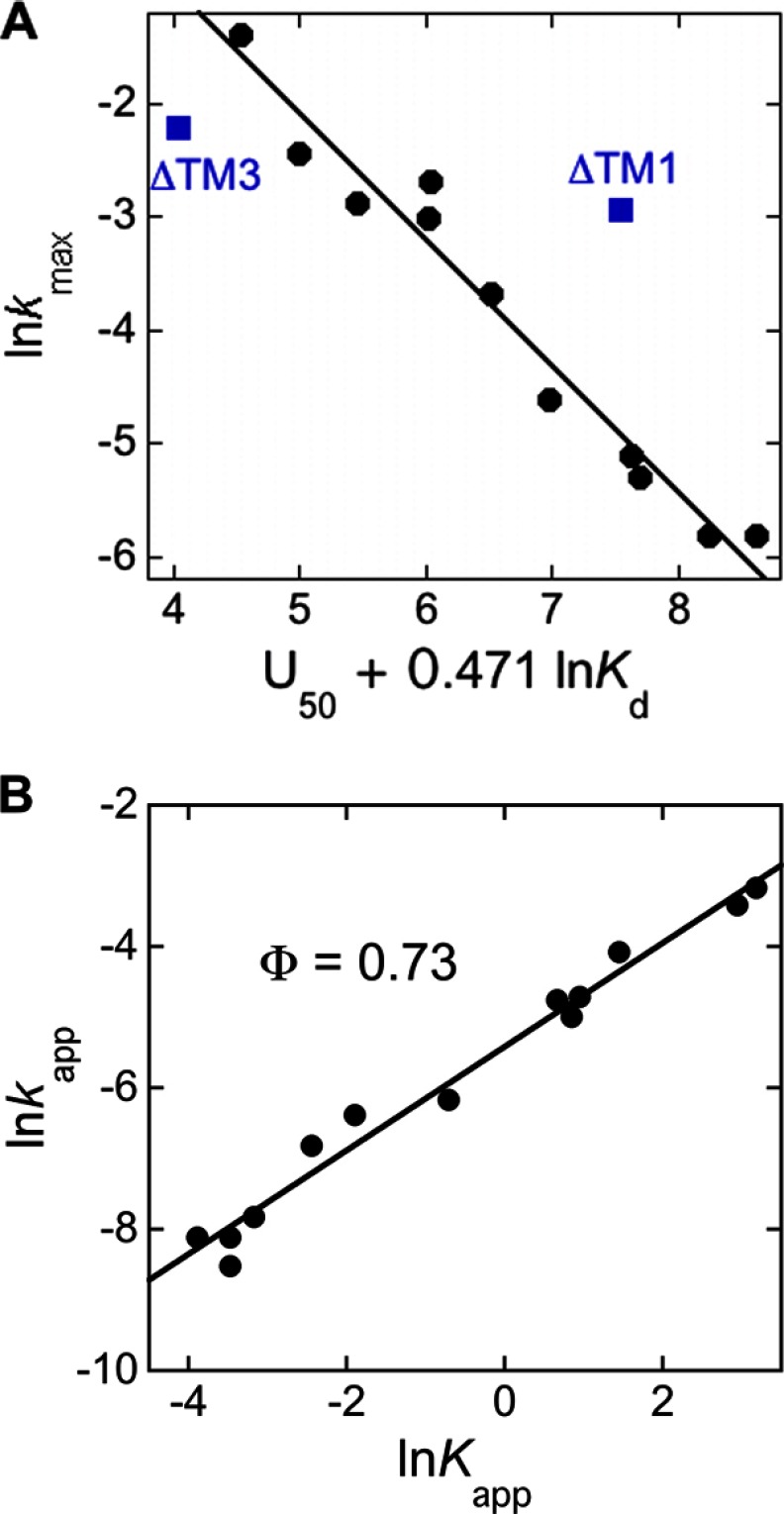
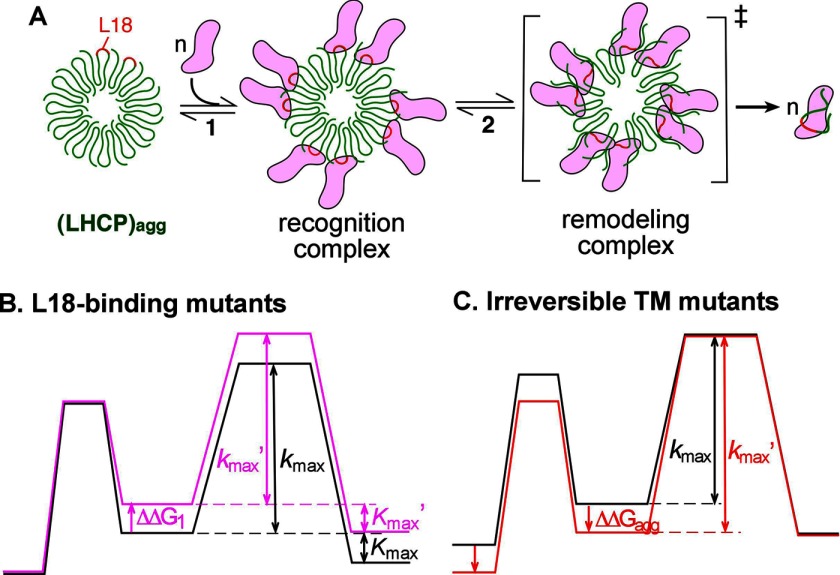
The ability of molecular chaperones to overcome the misfolding and aggregation of proteins is essential for the maintenance of proper protein homeostasis in all cells. Thus far, the best studied disaggregase systems are the Clp/Hsp100 family of "ATPases associated with various cellular activities" (AAA(+)) ATPases, which use mechanical forces powered by ATP hydrolysis to remodel protein aggregates. An alternative system to disassemble large protein aggregates is provided by the 38-kDa subunit of the chloroplast signal recognition particle (cpSRP43), which uses binding energy with its substrate proteins to drive disaggregation. The mechanism of this novel chaperone remains unclear. Here, molecular genetics and structure-activity analyses show that the action of cpSRP43 can be dissected into two steps with distinct molecular requirements: (i) initial recognition, during which cpSRP43 binds specifically to a recognition motif displayed on the surface of the aggregate; and (ii) aggregate remodeling, during which highly adaptable binding interactions of cpSRP43 with hydrophobic transmembrane domains of the substrate protein compete with the packing interactions within the aggregate. This establishes a useful framework to understand the molecular mechanism by which binding interactions from a molecular chaperone can be used to overcome protein aggregates in the absence of external energy input from ATP.
Keywords: ATP-independent Disaggregase; Enzyme Mechanisms; Kinetics; Membrane Proteins; Molecular Chaperone; Mutagenesis Mechanisms; Protein Aggregation; Protein Biogenesis; Signal Recognition Particle.
Figures
Similar articles
-
Conformational dynamics of a membrane protein chaperone enables spatially regulated substrate capture and release.Proc Natl Acad Sci U S A. 2016 Mar 22;113(12):E1615-24. doi: 10.1073/pnas.1524777113. Epub 2016 Mar 7. Proc Natl Acad Sci U S A. 2016. PMID: 26951662 Free PMC article.
-
Mechanism of an ATP-independent protein disaggregase: I. structure of a membrane protein aggregate reveals a mechanism of recognition by its chaperone.J Biol Chem. 2013 May 10;288(19):13420-30. doi: 10.1074/jbc.M113.462812. Epub 2013 Mar 22. J Biol Chem. 2013. PMID: 23525109 Free PMC article.
-
ATP-independent reversal of a membrane protein aggregate by a chloroplast SRP subunit.Nat Struct Mol Biol. 2010 Jun;17(6):696-702. doi: 10.1038/nsmb.1836. Epub 2010 Apr 27. Nat Struct Mol Biol. 2010. PMID: 20424608 Free PMC article.
-
Moonlighting chaperone-like activity of the universal regulatory 14-3-3 proteins.FEBS J. 2017 May;284(9):1279-1295. doi: 10.1111/febs.13986. Epub 2017 Jan 6. FEBS J. 2017. PMID: 27973707 Review.
-
Molecular mechanism of SRP-dependent light-harvesting protein transport to the thylakoid membrane in plants.Photosynth Res. 2018 Dec;138(3):303-313. doi: 10.1007/s11120-018-0544-6. Epub 2018 Jun 28. Photosynth Res. 2018. PMID: 29956039 Free PMC article. Review.
Cited by
-
Therapeutic Dissolution of Aberrant Phases by Nuclear-Import Receptors.Trends Cell Biol. 2019 Apr;29(4):308-322. doi: 10.1016/j.tcb.2018.12.004. Epub 2019 Jan 16. Trends Cell Biol. 2019. PMID: 30660504 Free PMC article. Review.
-
Two distinct sites of client protein interaction with the chaperone cpSRP43.J Biol Chem. 2018 Jun 8;293(23):8861-8873. doi: 10.1074/jbc.RA118.002215. Epub 2018 Apr 18. J Biol Chem. 2018. PMID: 29669809 Free PMC article.
-
An ankyrin repeat chaperone targets toxic oligomers during amyloidogenesis.Protein Sci. 2023 Aug;32(8):e4728. doi: 10.1002/pro.4728. Protein Sci. 2023. PMID: 37433015 Free PMC article.
-
Switchable client specificity in a dual functional chaperone coordinates light-harvesting complex biogenesis.Sci Adv. 2025 Jun 13;11(24):eadu5791. doi: 10.1126/sciadv.adu5791. Epub 2025 Jun 13. Sci Adv. 2025. PMID: 40512866 Free PMC article.
-
Conformational dynamics of a membrane protein chaperone enables spatially regulated substrate capture and release.Proc Natl Acad Sci U S A. 2016 Mar 22;113(12):E1615-24. doi: 10.1073/pnas.1524777113. Epub 2016 Mar 7. Proc Natl Acad Sci U S A. 2016. PMID: 26951662 Free PMC article.
References
-
- Balch W. E., Morimoto R. I., Dillin A., Kelly J. W. (2008) Adapting proteostasis for disease intervention. Science 319, 916–919 - PubMed
-
- Hartl F. U., Hayer-Hartl M. (2002) Molecular chaperones in the cytosol: from nascent chain to folded protein. Science 295, 1852–1858 - PubMed
-
- Chang H. C., Tang Y. C., Hayer-Hartl M., Hartl F. U. (2007) Snapshot: molecular chaperones, Part I. Cell 128, 212–213 - PubMed
-
- Tang Y. C., Chang H. C., Hayer-Hartl M., Hartl F. U. (2007) Snapshot: molecular chaperones, Part II. Cell 128, 412–413 - PubMed
-
- Doyle S. M., Wickner S. (2009) Hsp104 and ClpB: protein disaggregating machines. Trends Biochem. Sci. 34, 40–48 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
