

Geometric catalysis of membrane fission driven by flexible dynamin rings
- PMID: 23520112
- PMCID: PMC3980720
- DOI: 10.1126/science.1233920
Geometric catalysis of membrane fission driven by flexible dynamin rings
Abstract
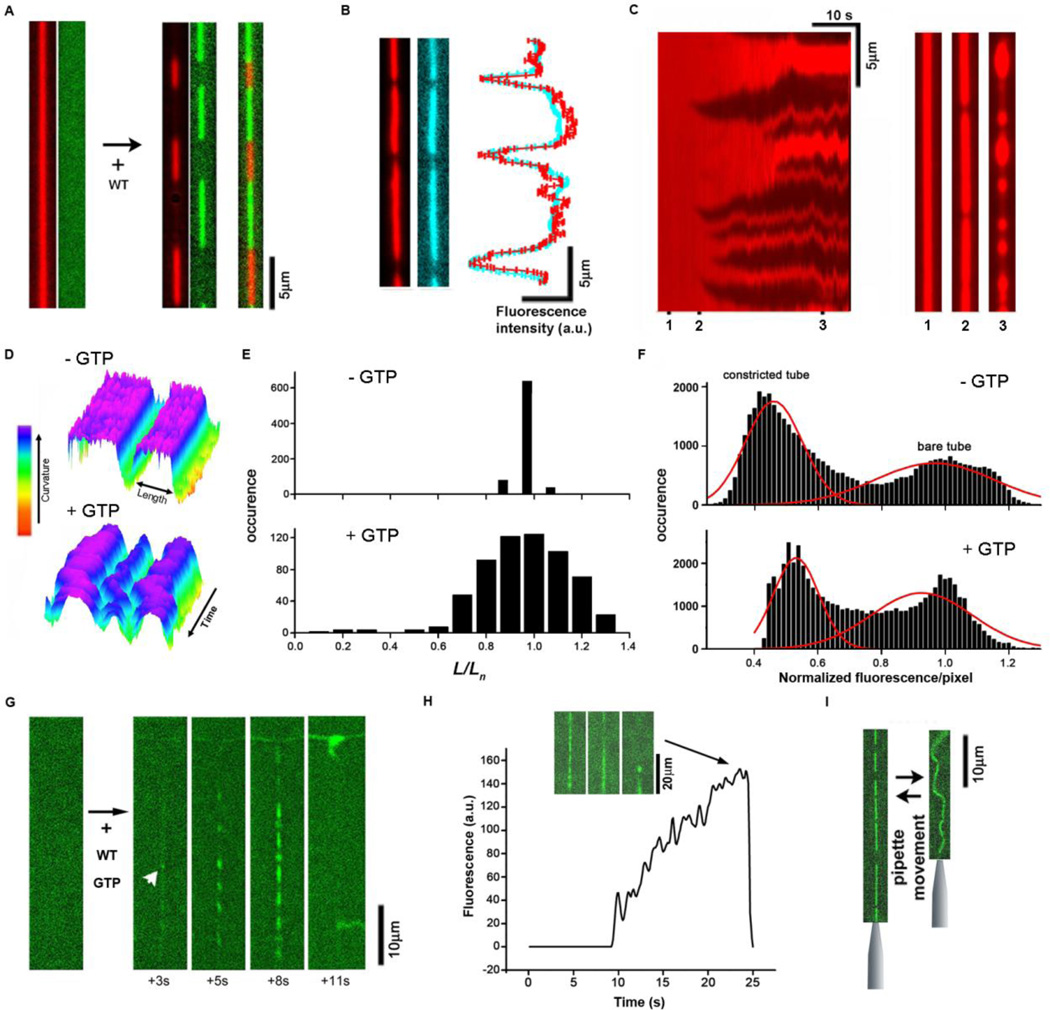
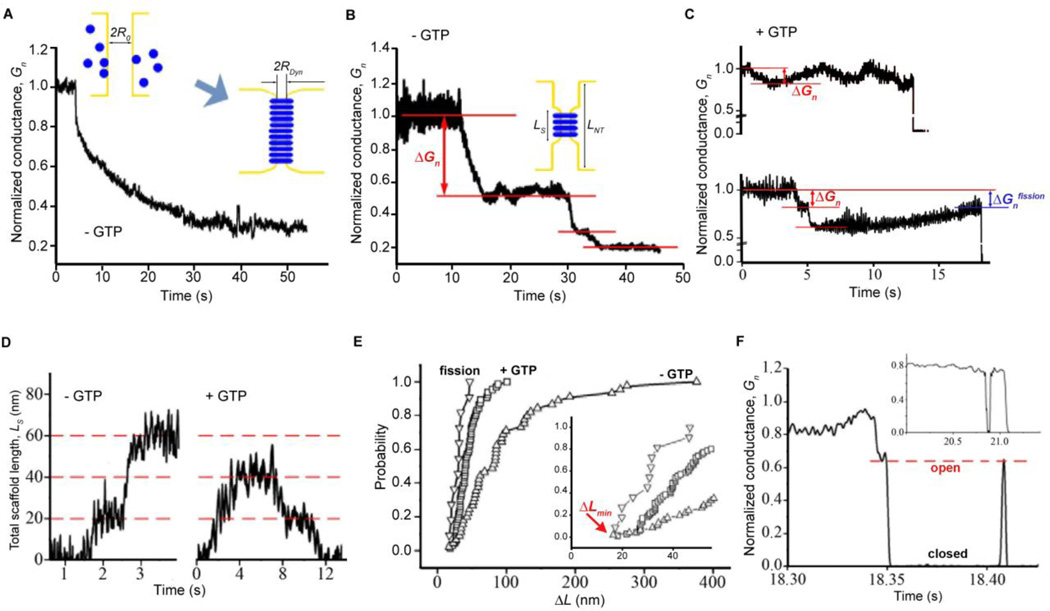
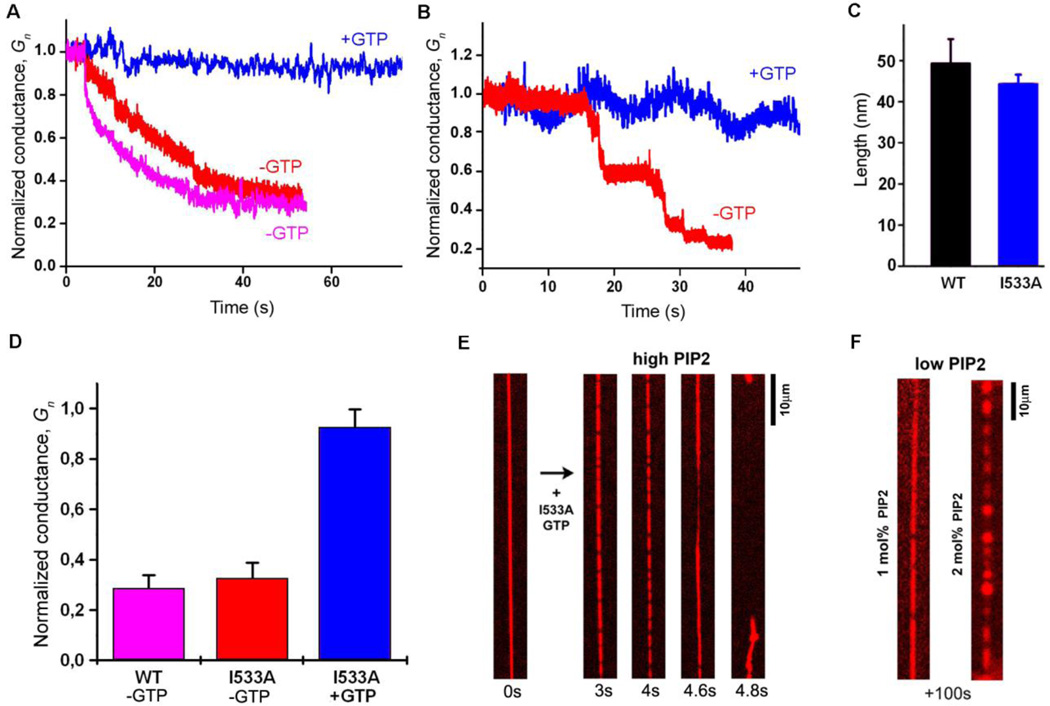
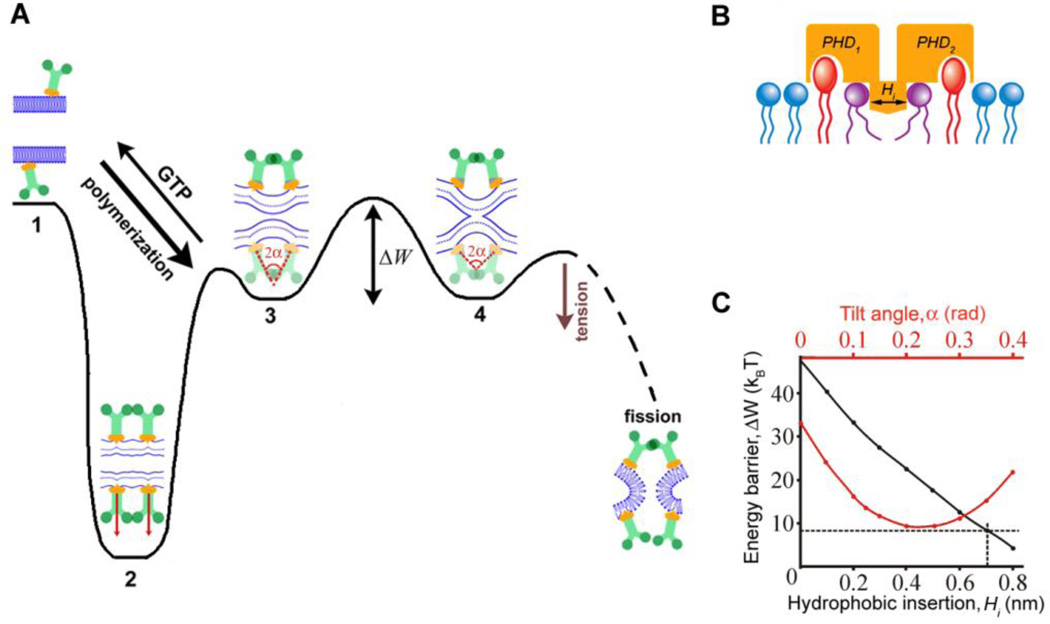
Biological membrane fission requires protein-driven stress. The guanosine triphosphatase (GTPase) dynamin builds up membrane stress by polymerizing into a helical collar that constricts the neck of budding vesicles. How this curvature stress mediates nonleaky membrane remodeling is actively debated. Using lipid nanotubes as substrates to directly measure geometric intermediates of the fission pathway, we found that GTP hydrolysis limits dynamin polymerization into short, metastable collars that are optimal for fission. Collars as short as two rungs translated radial constriction to reversible hemifission via membrane wedging of the pleckstrin homology domains (PHDs) of dynamin. Modeling revealed that tilting of the PHDs to conform with membrane deformations creates the low-energy pathway for hemifission. This local coordination of dynamin and lipids suggests how membranes can be remodeled in cells.
Figures
Comment in
-
Cell biology. Dynamin flexibility drives fission.Science. 2013 Mar 22;339(6126):1392-3. doi: 10.1126/science.1236005. Science. 2013. PMID: 23520100 No abstract available.
Similar articles
-
Cryo-EM of the dynamin polymer assembled on lipid membrane.Nature. 2018 Aug;560(7717):258-262. doi: 10.1038/s41586-018-0378-6. Epub 2018 Aug 1. Nature. 2018. PMID: 30069048 Free PMC article.
-
Flexible pivoting of dynamin pleckstrin homology domain catalyzes fission: insights into molecular degrees of freedom.Mol Biol Cell. 2021 Jul 1;32(14):1306-1319. doi: 10.1091/mbc.E20-12-0794. Epub 2021 May 12. Mol Biol Cell. 2021. PMID: 33979205 Free PMC article.
-
Dynamin: functional design of a membrane fission catalyst.Annu Rev Cell Dev Biol. 2011;27:79-105. doi: 10.1146/annurev-cellbio-100109-104016. Epub 2011 May 18. Annu Rev Cell Dev Biol. 2011. PMID: 21599493 Review.
-
GTPase cycle of dynamin is coupled to membrane squeeze and release, leading to spontaneous fission.Cell. 2008 Dec 26;135(7):1276-86. doi: 10.1016/j.cell.2008.11.028. Epub 2008 Dec 11. Cell. 2008. PMID: 19084269 Free PMC article.
-
Building a fission machine--structural insights into dynamin assembly and activation.J Cell Sci. 2013 Jul 1;126(Pt 13):2773-84. doi: 10.1242/jcs.108845. Epub 2013 Jun 18. J Cell Sci. 2013. PMID: 23781021 Free PMC article. Review.
Cited by
-
Intravesicular Solute Delivery and Surface Area Regulation in Giant Unilamellar Vesicles Driven by Cycles of Osmotic Stresses.J Am Chem Soc. 2024 Feb 7;146(5):3250-3261. doi: 10.1021/jacs.3c11679. Epub 2024 Jan 24. J Am Chem Soc. 2024. PMID: 38266489 Free PMC article.
-
Reducing dynamin 2 expression rescues X-linked centronuclear myopathy.J Clin Invest. 2014 Mar;124(3):1350-63. doi: 10.1172/JCI71206. Epub 2014 Feb 24. J Clin Invest. 2014. PMID: 24569376 Free PMC article.
-
Combining patch-clamping and fluorescence microscopy for quantitative reconstitution of cellular membrane processes with Giant Suspended Bilayers.Sci Rep. 2019 May 10;9(1):7255. doi: 10.1038/s41598-019-43561-4. Sci Rep. 2019. PMID: 31076583 Free PMC article.
-
Reaching a consensus on the mechanism of dynamin?F1000Prime Rep. 2014 Oct 1;6:86. doi: 10.12703/P6-86. eCollection 2014. F1000Prime Rep. 2014. PMID: 25374664 Free PMC article. Review.
-
Pore formation in lipid membrane I: Continuous reversible trajectory from intact bilayer through hydrophobic defect to transversal pore.Sci Rep. 2017 Sep 22;7(1):12152. doi: 10.1038/s41598-017-12127-7. Sci Rep. 2017. PMID: 28939906 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
