

Renal hypodysplasia associates with a WNT4 variant that causes aberrant canonical WNT signaling
- PMID: 23520208
- PMCID: PMC3609130
- DOI: 10.1681/ASN.2012010097
Renal hypodysplasia associates with a WNT4 variant that causes aberrant canonical WNT signaling
Abstract
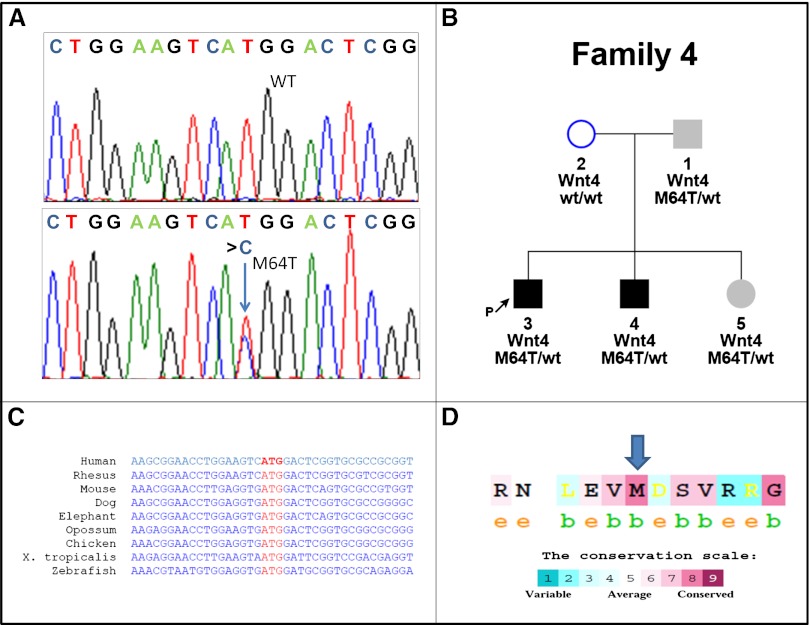
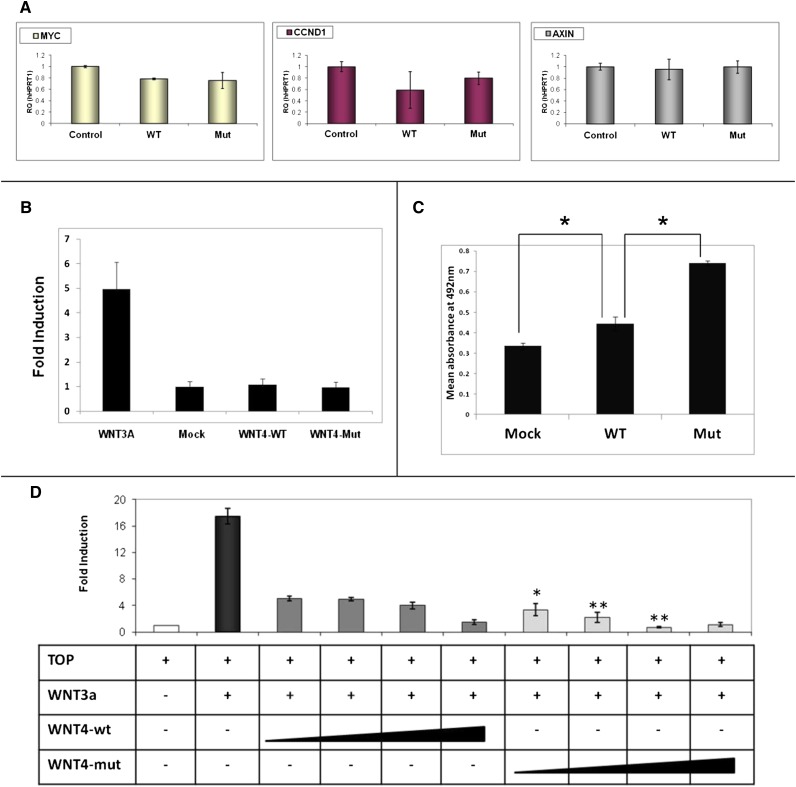
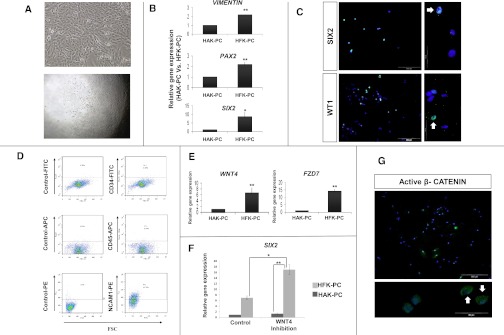
Abnormal differentiation of the renal stem/progenitor pool into kidney tissue can lead to renal hypodysplasia (RHD), but the underlying causes of RHD are not well understood. In this multicenter study, we identified 20 Israeli pedigrees with isolated familial, nonsyndromic RHD and screened for mutations in candidate genes involved in kidney development, including PAX2, HNF1B, EYA1, SIX1, SIX2, SALL1, GDNF, WNT4, and WT1. In addition to previously reported RHD-causing genes, we found that two affected brothers were heterozygous for a missense variant in the WNT4 gene. Functional analysis of this variant revealed both antagonistic and agonistic canonical WNT stimuli, dependent on cell type. In HEK293 cells, WNT4 inhibited WNT3A induced canonical activation, and the WNT4 variant significantly enhanced this inhibition of the canonical WNT pathway. In contrast, in primary cultures of human fetal kidney cells, which maintain WNT activation and more closely represent WNT signaling in renal progenitors during nephrogenesis, this mutation caused significant loss of function, resulting in diminished canonical WNT/β-catenin signaling. In conclusion, heterozygous WNT4 variants are likely to play a causative role in renal hypodysplasia.
Figures

Similar articles
-
Prevalence of mutations in renal developmental genes in children with renal hypodysplasia: results of the ESCAPE study.J Am Soc Nephrol. 2006 Oct;17(10):2864-70. doi: 10.1681/ASN.2006030277. Epub 2006 Sep 13. J Am Soc Nephrol. 2006. PMID: 16971658
-
Transcriptional regulatory control of mammalian nephron progenitors revealed by multi-factor cistromic analysis and genetic studies.PLoS Genet. 2018 Jan 29;14(1):e1007181. doi: 10.1371/journal.pgen.1007181. eCollection 2018 Jan. PLoS Genet. 2018. PMID: 29377931 Free PMC article.
-
Restoration of WNT4 inhibits cell growth in leukemia-derived cell lines.BMC Cancer. 2013 Nov 25;13:557. doi: 10.1186/1471-2407-13-557. BMC Cancer. 2013. PMID: 24274766 Free PMC article.
-
WNT4 Balances Development vs Disease in Gynecologic Tissues and Women's Health.Endocrinology. 2021 Jul 1;162(7):bqab093. doi: 10.1210/endocr/bqab093. Endocrinology. 2021. PMID: 33963381 Free PMC article. Review.
-
Roles and action mechanisms of WNT4 in cell differentiation and human diseases: a review.Cell Death Discov. 2021 Oct 12;7(1):287. doi: 10.1038/s41420-021-00668-w. Cell Death Discov. 2021. PMID: 34642299 Free PMC article. Review.
Cited by
-
From skeletal to cardiovascular disease in 12 steps-the evolution of sclerostin as a major player in CKD-MBD.Pediatr Nephrol. 2016 Feb;31(2):195-206. doi: 10.1007/s00467-015-3069-7. Epub 2015 Mar 4. Pediatr Nephrol. 2016. PMID: 25735207 Review.
-
A human integrin-α3 mutation confers major renal developmental defects.PLoS One. 2014 Mar 12;9(3):e90879. doi: 10.1371/journal.pone.0090879. eCollection 2014. PLoS One. 2014. PMID: 24621570 Free PMC article.
-
Next-generation sequencing for research and diagnostics in kidney disease.Nat Rev Nephrol. 2014 Aug;10(8):433-44. doi: 10.1038/nrneph.2014.95. Epub 2014 Jun 10. Nat Rev Nephrol. 2014. PMID: 24914583 Review.
-
Developmental Genetics and Congenital Anomalies of the Kidney and Urinary Tract.J Pediatr Genet. 2016 Mar;5(1):51-60. doi: 10.1055/s-0035-1558423. Epub 2015 Sep 7. J Pediatr Genet. 2016. PMID: 27617142 Free PMC article. Review.
-
Modeling congenital kidney diseases in Xenopus laevis.Dis Model Mech. 2019 Apr 9;12(4):dmm038604. doi: 10.1242/dmm.038604. Dis Model Mech. 2019. PMID: 30967415 Free PMC article. Review.
References
-
- Weber S, Moriniere V, Knüppel T, Charbit M, Dusek J, Ghiggeri GM, Jankauskiené A, Mir S, Montini G, Peco-Antic A, Wühl E, Zurowska AM, Mehls O, Antignac C, Schaefer F, Salomon R: Prevalence of mutations in renal developmental genes in children with renal hypodysplasia: Results of the ESCAPE study. J Am Soc Nephrol 17: 2864–2870, 2006 - PubMed
-
- Eccles MR, Schimmenti LA: Renal-coloboma syndrome: A multi-system developmental disorder caused by PAX2 mutations. Clin Genet 56: 1–9, 1999 - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
