

Depletion of cellular iron by curcumin leads to alteration in histone acetylation and degradation of Sml1p in Saccharomyces cerevisiae
- PMID: 23520547
- PMCID: PMC3592818
- DOI: 10.1371/journal.pone.0059003
Depletion of cellular iron by curcumin leads to alteration in histone acetylation and degradation of Sml1p in Saccharomyces cerevisiae
Erratum in
- PLoS One. 2013;8(10). doi:10.1371/annotation/ac974676-18a6-4aee-b38e-57a1142a3dc1
Abstract
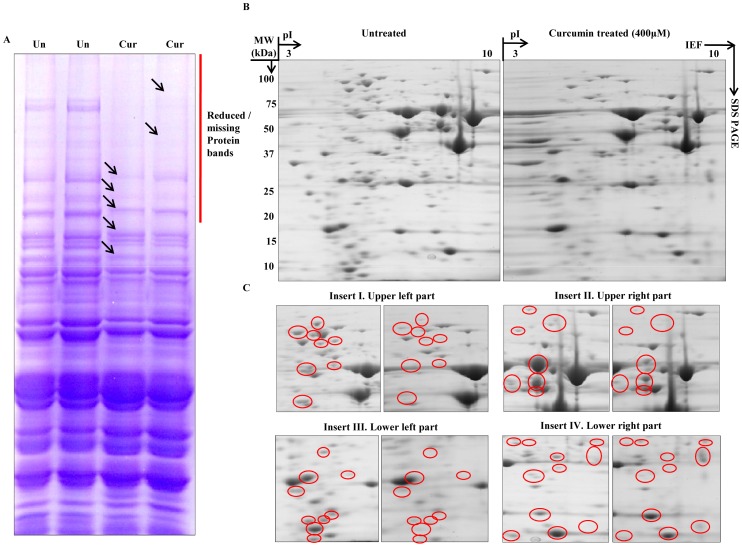
Curcumin, a naturally occurring polyphenolic compound, is known to possess diverse pharmacological properties. There is a scarcity of literature documenting the exact mechanism by which curcumin modulates its biological effects. In the present study, we have used yeast as a model organism to dissect the mechanism underlying the action of curcumin. We found that the yeast mutants of histone proteins and chromatin modifying enzymes were sensitive to curcumin and further supplementation of iron resulted in reversal of the changes induced by curcumin. Additionally, treatment of curcumin caused the iron starvation induced expression of FET3, FRE1 genes. We also demonstrated that curcumin induces degradation of Sml1p, a ribonucleotide reductase inhibitor involved in regulating dNTPs production. The degradation of Sml1p was mediated through proteasome and vacuole dependent protein degradation pathways. Furthermore, curcumin exerts biological effect by altering global proteome profile without affecting chromatin architecture. These findings suggest that the medicinal properties of curcumin are largely contributed by its cumulative effect of iron starvation and epigenetic modifications.
Conflict of interest statement
Figures







References
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ (1997) Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389: 251–260. - PubMed
-
- Clapier CR, Cairns BR (2009) The biology of chromatin remodeling complexes. Annu Rev Biochem 78: 273–304. - PubMed
-
- Huertas D, Sendra R, Munoz P (2009) Chromatin dynamics coupled to DNA repair. Epigenetics 4: 31–42. - PubMed
-
- Ehrenhofer-Murray AE (2004) Chromatin dynamics at DNA replication, transcription and repair. Eur J Biochem 271: 2335–2349. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
