

ER-PM connections: sites of information transfer and inter-organelle communication
- PMID: 23522446
- PMCID: PMC4074705
- DOI: 10.1016/j.ceb.2013.02.020
ER-PM connections: sites of information transfer and inter-organelle communication
Abstract
Eukaryotic cells are divided into distinct membrane-bound organelles with unique identities and specialized metabolic functions. Communication between organelles must take place to regulate the size, shape, and composition of individual organelles, as well as to coordinate transport between organelles. The endoplasmic reticulum (ER) forms an expansive membrane network that contacts and participates in crosstalk with several other organelles in the cell, most notably the plasma membrane (PM). ER-PM junctions have well-established functions in the movement of small molecules, such as lipids and ions, between the ER and PM. Recent discoveries have revealed additional exciting roles for ER-PM junctions in the regulation of cell signaling, ER shape and architecture, and PM domain organization.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
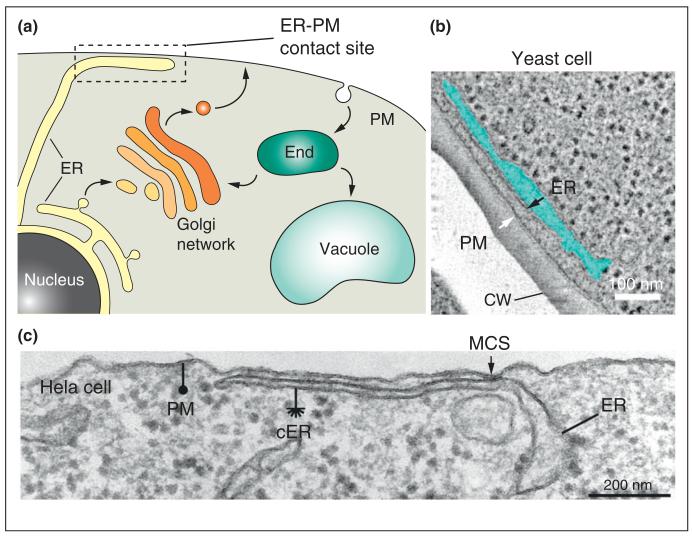
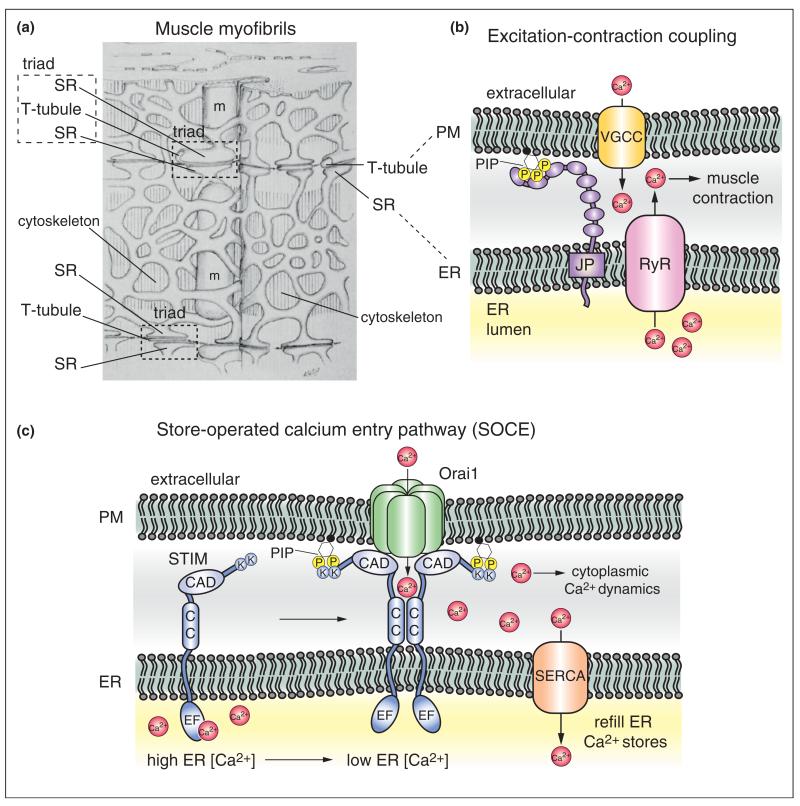
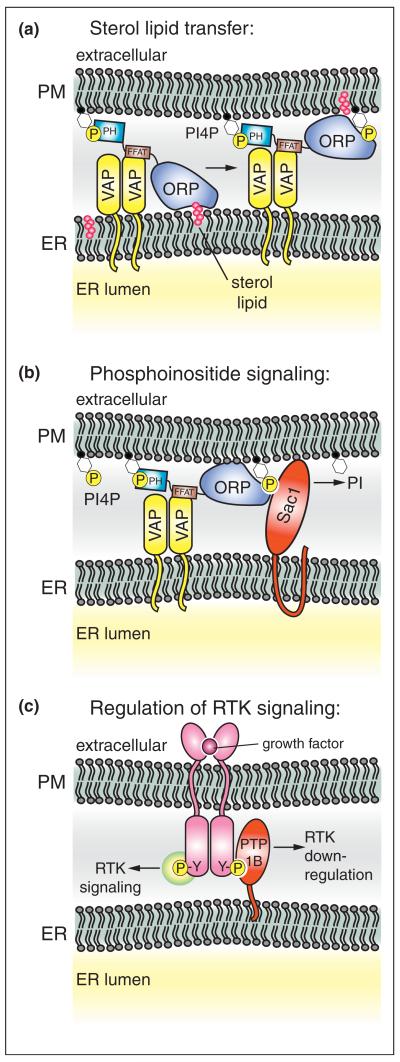
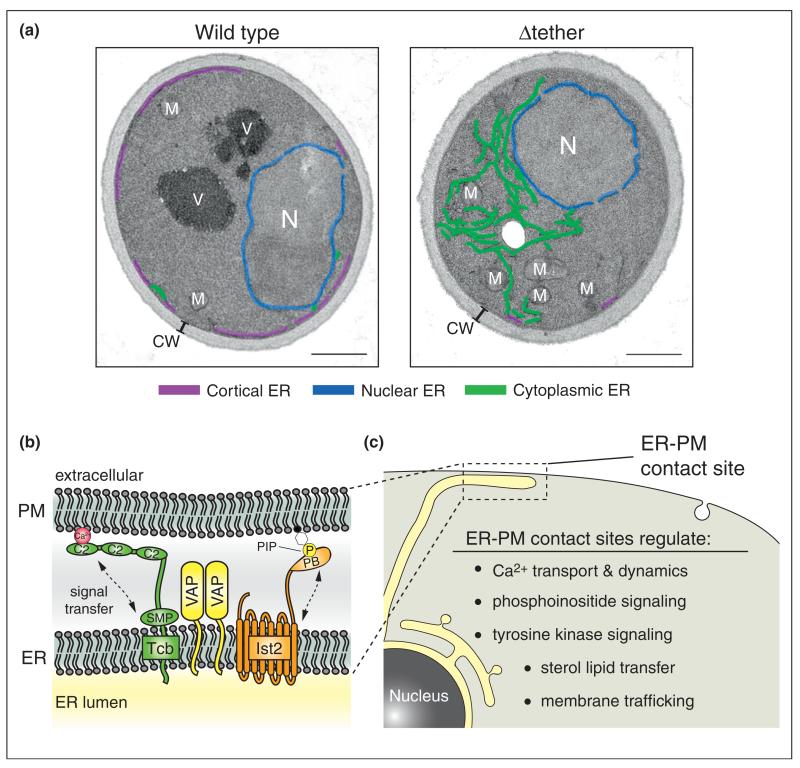
References
-
- Elbaz Y, Schuldiner M. Staying in touch: the molecular era of organelle contact sites. Trends Biochem Sci. 2011;36:616–623. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
