

Signaling pathways differentially affect RNA polymerase II initiation, pausing, and elongation rate in cells
- PMID: 23523369
- PMCID: PMC3640649
- DOI: 10.1016/j.molcel.2013.02.015
Signaling pathways differentially affect RNA polymerase II initiation, pausing, and elongation rate in cells
Erratum in
- Mol Cell. 2013 Jun 6;50(5):778
Abstract
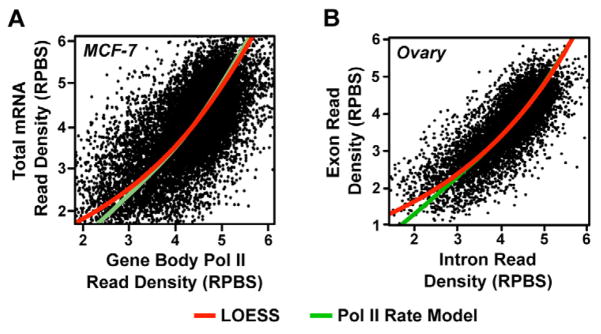
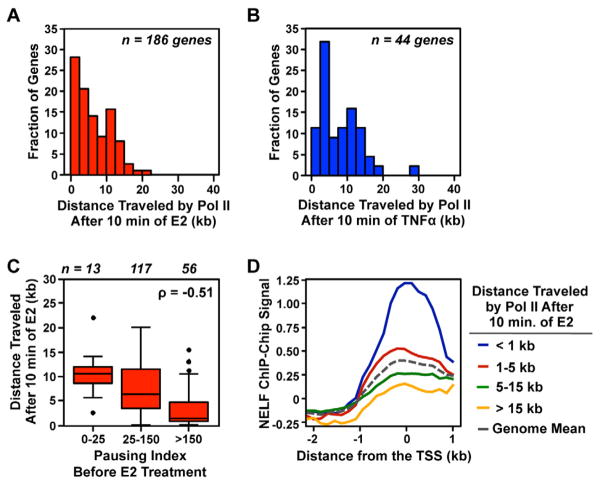
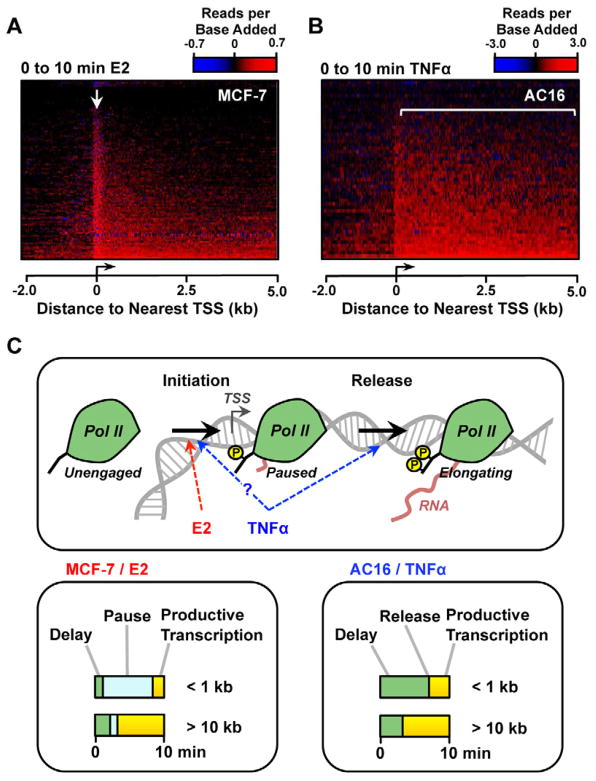
RNA polymerase II (Pol II) transcribes hundreds of kilobases of DNA, limiting the production of mRNAs and lncRNAs. We used global run-on sequencing (GRO-seq) to measure the rates of transcription by Pol II following gene activation. Elongation rates vary as much as 4-fold at different genomic loci and in response to two distinct cellular signaling pathways (i.e., 17β-estradiol [E2] and TNF-α). The rates are slowest near the promoter and increase during the first ~15 kb transcribed. Gene body elongation rates correlate with Pol II density, resulting in systematically higher rates of transcript production at genes with higher Pol II density. Pol II dynamics following short inductions indicate that E2 stimulates gene expression by increasing Pol II initiation, whereas TNF-α reduces Pol II residence time at pause sites. Collectively, our results identify previously uncharacterized variation in the rate of transcription and highlight elongation as an important, variable, and regulated rate-limiting step during transcription.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
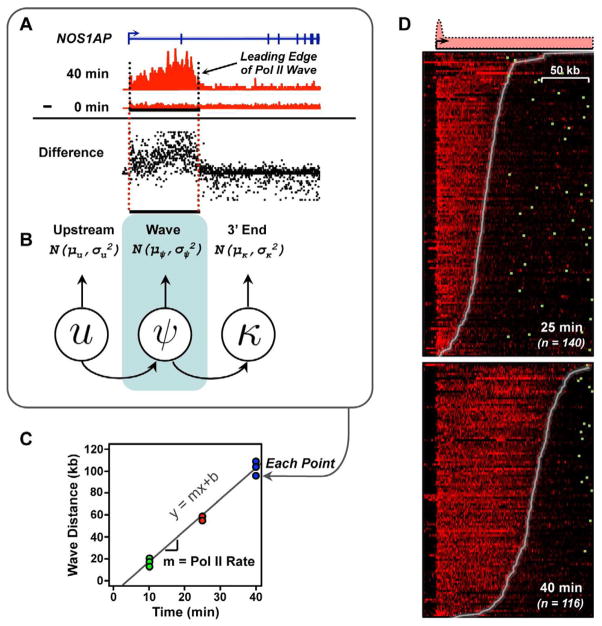
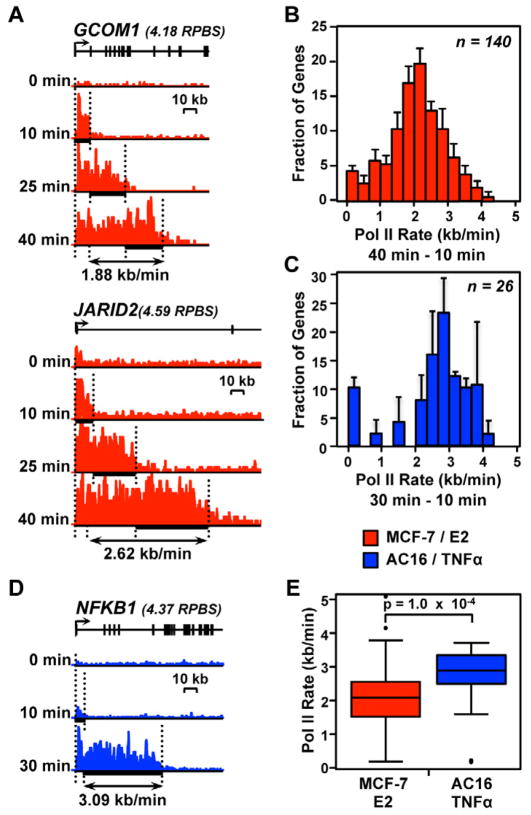
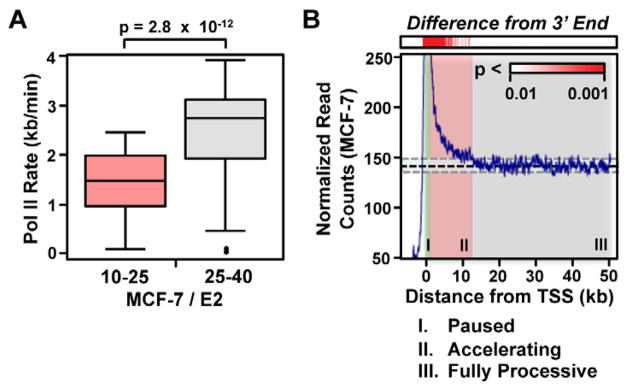
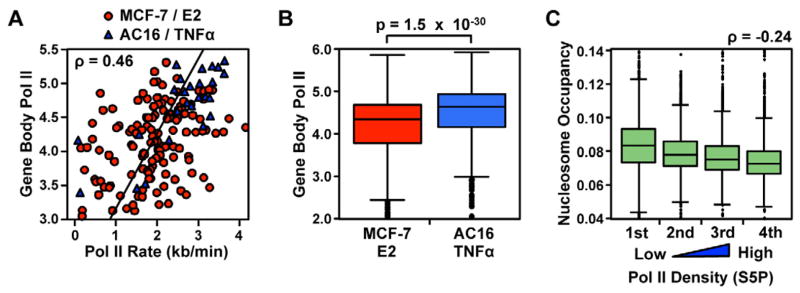
Comment in
-
Catching the waves: following the leading edge of elongating RNA polymerase II.Mol Cell. 2013 Apr 25;50(2):159-60. doi: 10.1016/j.molcel.2013.04.009. Mol Cell. 2013. PMID: 23622514 Free PMC article.
References
-
- Ardehali MB, Lis JT. Tracking rates of transcription and splicing in vivo. Nat Struct Mol Biol. 2009;16:1123–1124. - PubMed
-
- Barboric M, Nissen RM, Kanazawa S, Jabrane-Ferrat N, Peterlin BM. NF-kappaB binds P-TEFb to stimulate transcriptional elongation by RNA polymerase II. Mol Cell. 2001;8:327–337. - PubMed
-
- Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–837. - PubMed
-
- Belotserkovskaya R, Oh S, Bondarenko VA, Orphanides G, Studitsky VM, Reinberg D. FACT facilitates transcription-dependent nucleosome alteration. Science. 2003;301:1090–1093. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
