

Emerging roles for chromatin as a signal integration and storage platform
- PMID: 23524488
- PMCID: PMC4082330
- DOI: 10.1038/nrm3545
Emerging roles for chromatin as a signal integration and storage platform
Abstract
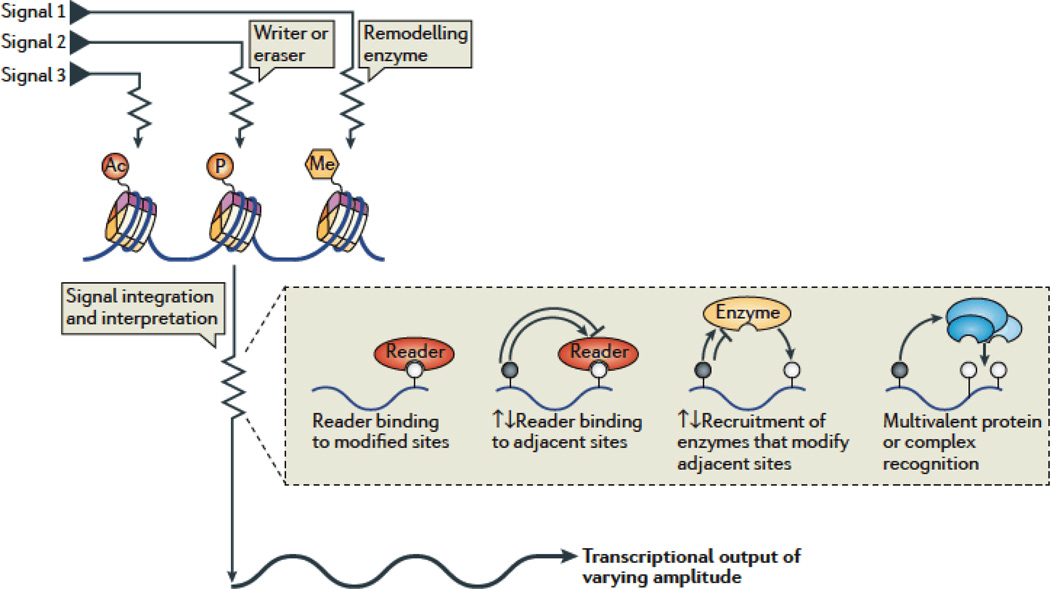
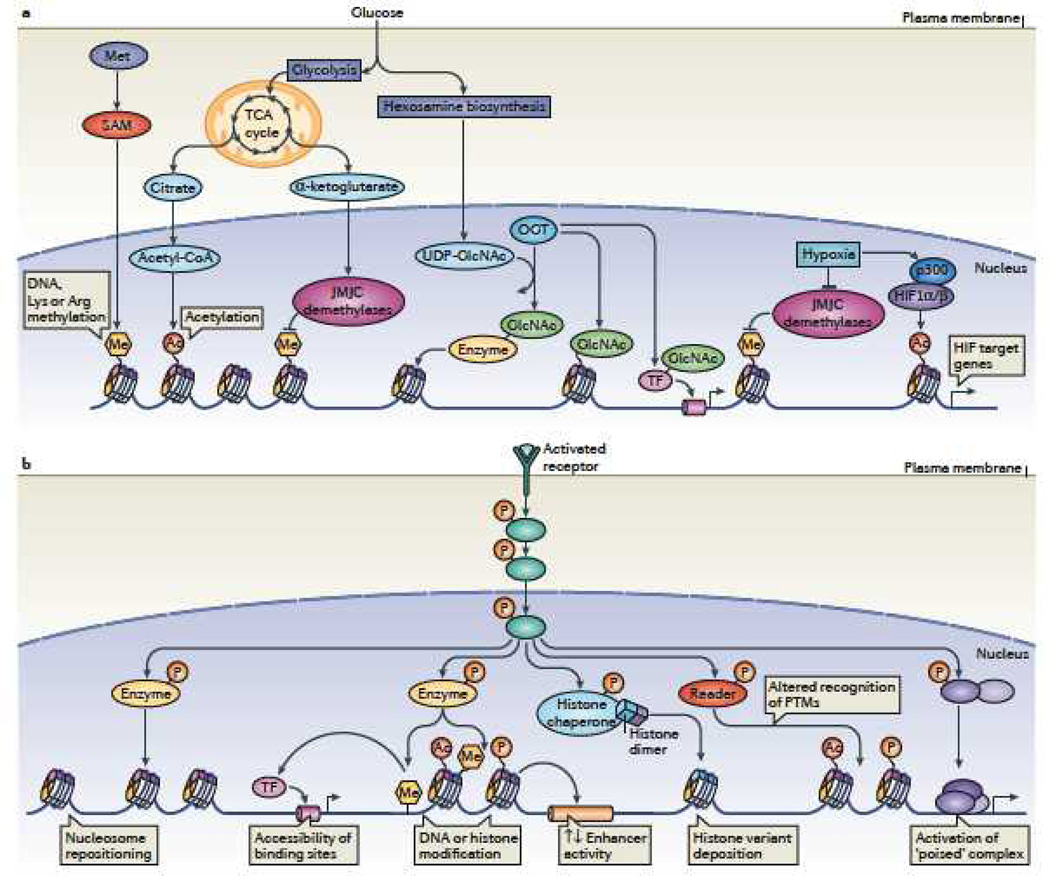
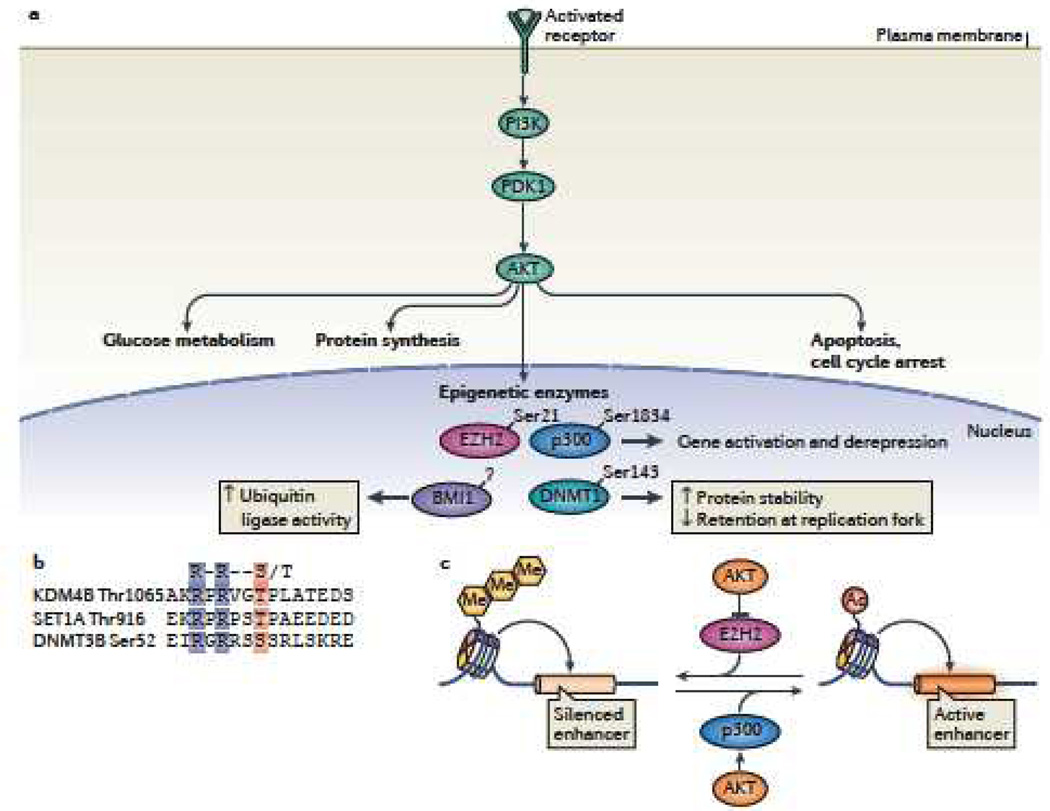
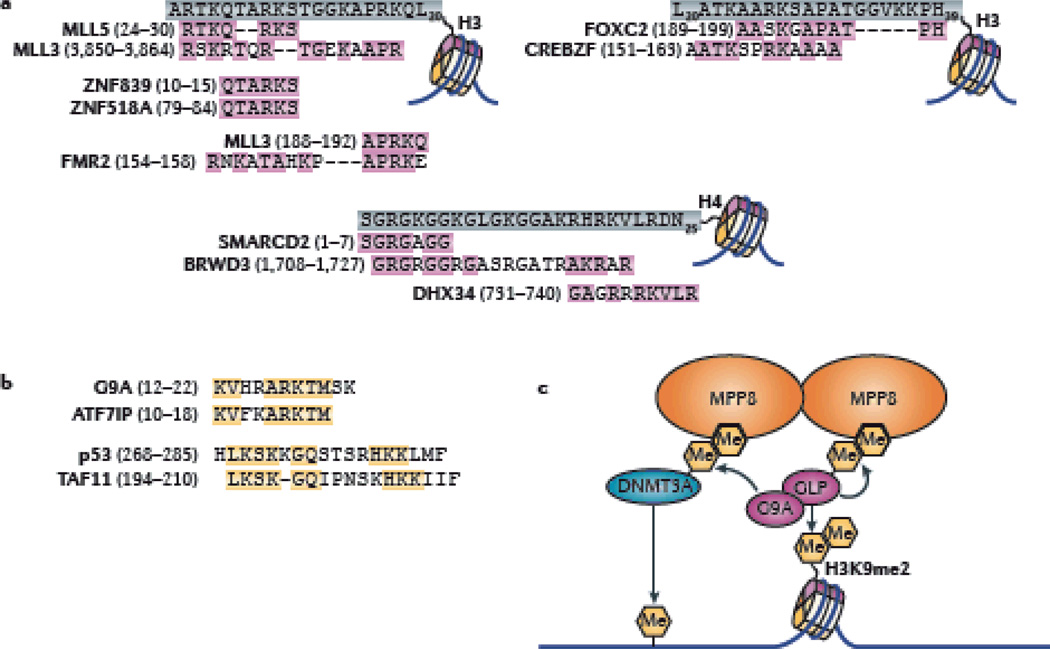
Cells of a multicellular organism, all containing nearly identical genetic information, respond to differentiation cues in variable ways. In addition, cells are plastic, able to execute their specialized function while maintaining the ability to adapt to environmental changes. This is achieved through multiple mechanisms, including the direct regulation of chromatin-based processes in response to stimuli. How signal transduction pathways directly communicate with chromatin to change the epigenetic landscape is poorly understood. The preponderance of covalent modifications on histone tails coupled with a relatively small number of functional outputs raises the possibility that chromatin acts as a site of signal integration and storage.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
