

OTUB1 modulates c-IAP1 stability to regulate signalling pathways
- PMID: 23524849
- PMCID: PMC3630360
- DOI: 10.1038/emboj.2013.62
OTUB1 modulates c-IAP1 stability to regulate signalling pathways
Abstract
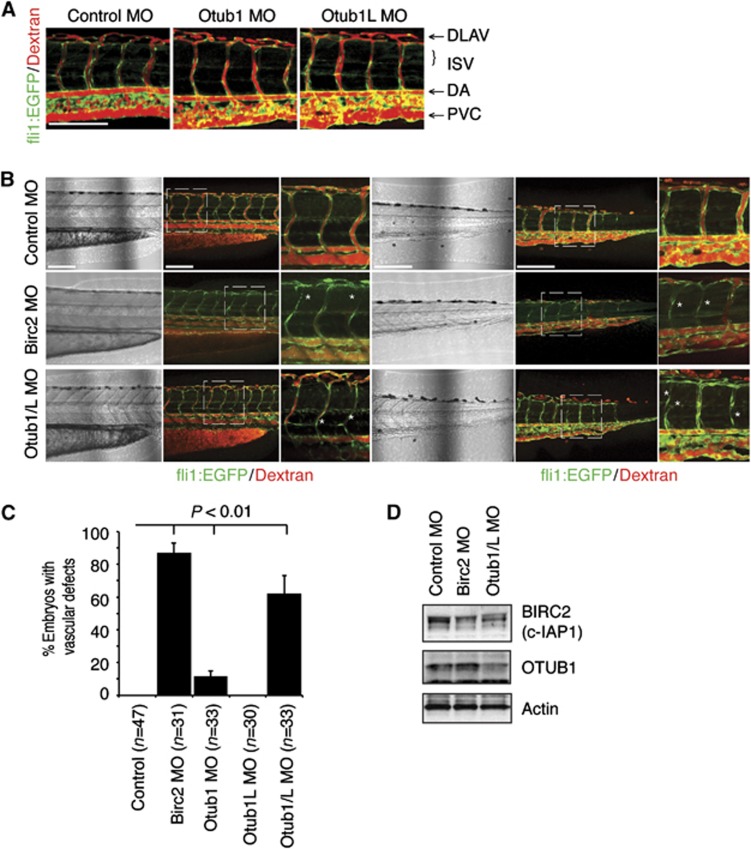
The cellular inhibitor of apoptosis (c-IAP) proteins are E3 ubiquitin ligases that are critical regulators of tumour necrosis factor (TNF) receptor (TNFR)-mediated signalling. Through their E3 ligase activity c-IAP proteins promote ubiquitination of receptor-interaction protein 1 (RIP1), NF-κB-inducing kinase (NIK) and themselves, and regulate the assembly of TNFR signalling complexes. Consequently, in the absence of c-IAP proteins, TNFR-mediated activation of NF-κB and MAPK pathways and the induction of gene expression are severely reduced. Here, we describe the identification of OTUB1 as a c-IAP-associated deubiquitinating enzyme that regulates c-IAP1 stability. OTUB1 disassembles K48-linked polyubiquitin chains from c-IAP1 in vitro and in vivo within the TWEAK receptor-signalling complex. Downregulation of OTUB1 promotes TWEAK- and IAP antagonist-stimulated caspase activation and cell death, and enhances c-IAP1 degradation. Furthermore, knockdown of OTUB1 reduces TWEAK-induced activation of canonical NF-κB and MAPK signalling pathways and modulates TWEAK-induced gene expression. Finally, suppression of OTUB1 expression in zebrafish destabilizes c-IAP (Birc2) protein levels and disrupts fish vasculature. These results suggest that OTUB1 regulates NF-κB and MAPK signalling pathways and TNF-dependent cell death by modulating c-IAP1 stability.
Conflict of interest statement
All of the authors are employees of Genentech, Inc.
Figures
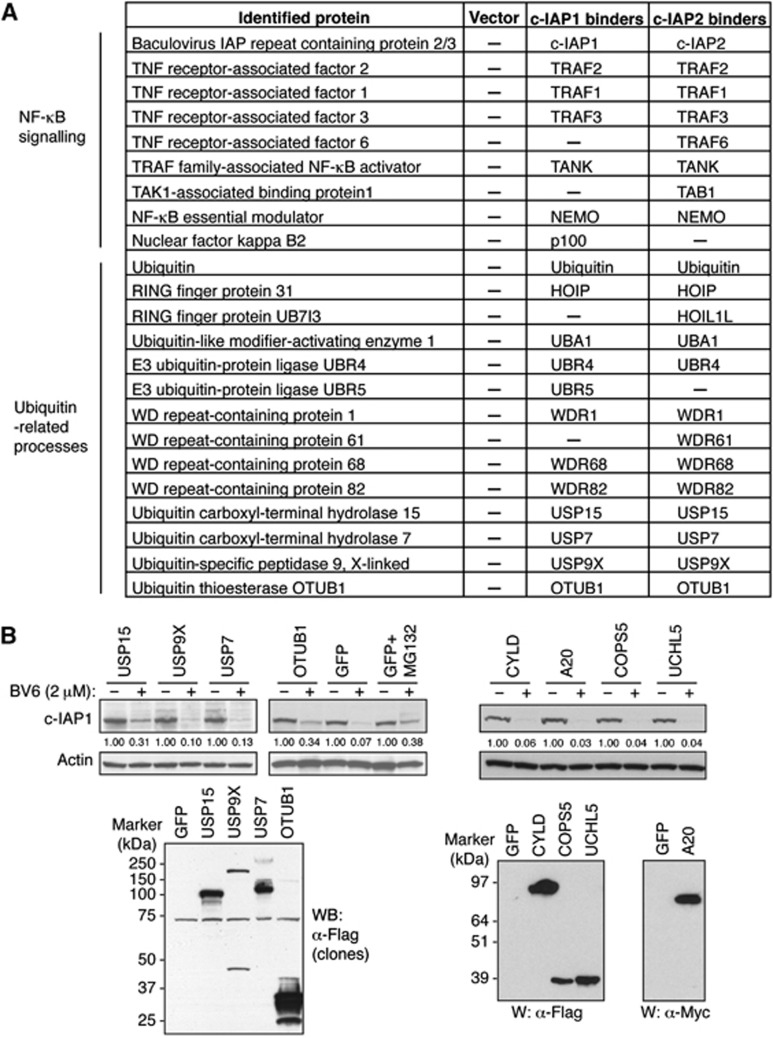
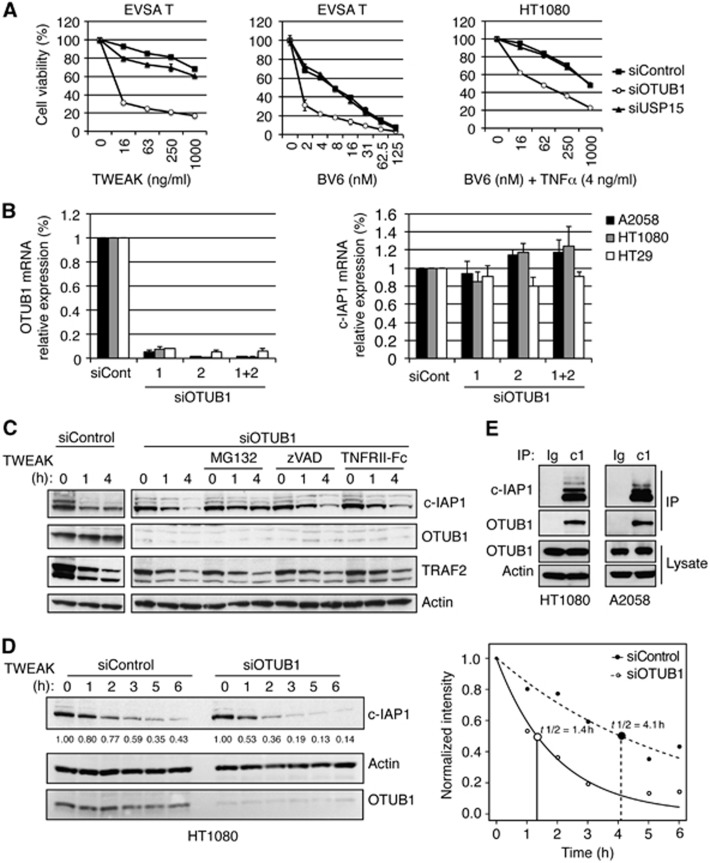

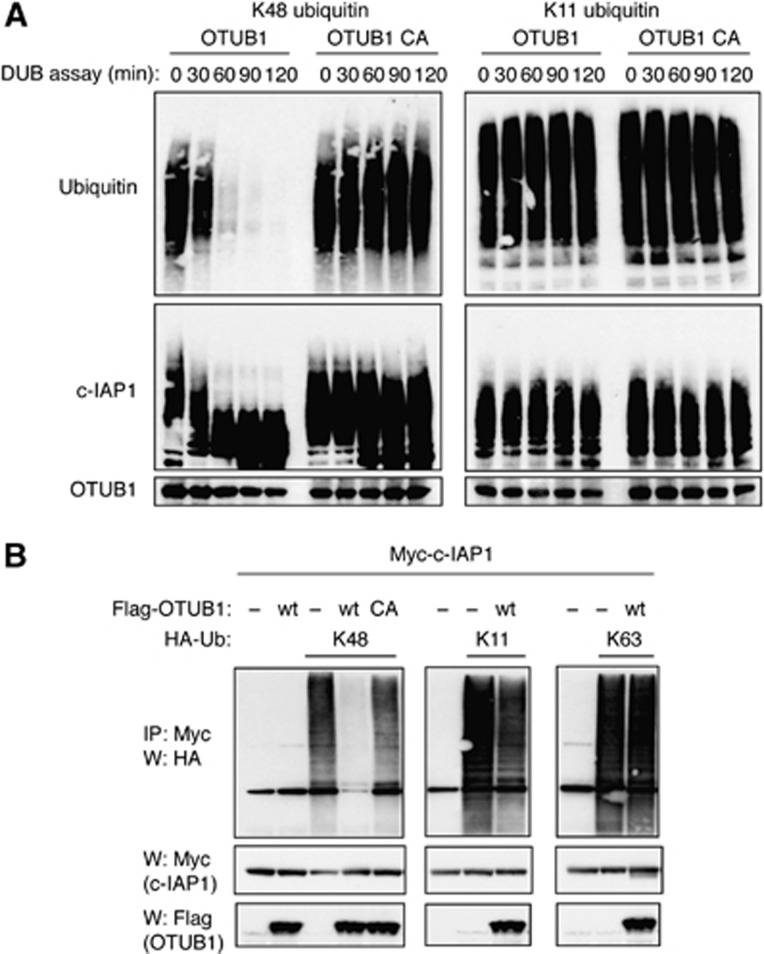

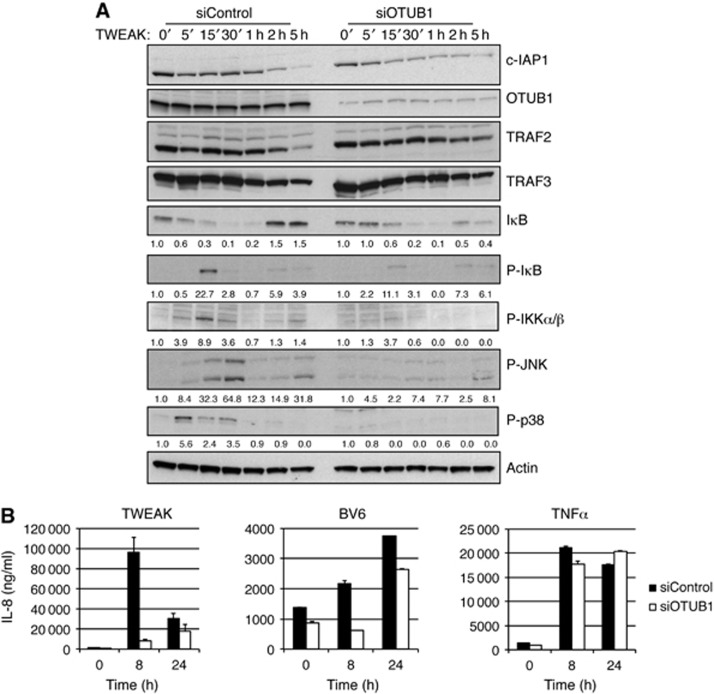
References
-
- Annunziata CM, Davis RE, Demchenko Y, Bellamy W, Gabrea A, Zhan F, Lenz G, Hanamura I, Wright G, Xiao W, Dave S, Hurt EM, Tan B, Zhao H, Stephens O, Santra M, Williams DR, Dang L, Barlogie B, Shaughnessy JD Jr. et al. (2007) Frequent engagement of the classical and alternative NF-kappaB pathways by diverse genetic abnormalities in multiple myeloma. Cancer Cell 12: 115–130 - PMC - PubMed
-
- Bertrand MJ, Milutinovic S, Dickson KM, Ho WC, Boudreault A, Durkin J, Gillard JW, Jaquith JB, Morris SJ, Barker PA (2008) cIAP1 and cIAP2 facilitate cancer cell survival by functioning as E3 ligases that promote RIP1 ubiquitination. Mol Cell 30: 689–700 - PubMed
-
- Blankenship JW, Varfolomeev E, Goncharov T, Fedorova AV, Kirkpatrick DS, Izrael-Tomasevic A, Phu L, Arnott D, Aghajan M, Zobel K, Bazan JF, Fairbrother WJ, Deshayes K, Vucic D (2009) Ubiquitin binding modulates IAP antagonist-stimulated proteasomal degradation of c-IAP1 and c-IAP2. Biochem J 417: 149–160 - PubMed
-
- Deshaies RJ, Joazeiro CA (2009) RING domain E3 ubiquitin ligases. Annu Rev Biochem 78: 399–434 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
