

The transcriptional response of Cryptococcus neoformans to ingestion by Acanthamoeba castellanii and macrophages provides insights into the evolutionary adaptation to the mammalian host
- PMID: 23524994
- PMCID: PMC3647765
- DOI: 10.1128/EC.00073-13
The transcriptional response of Cryptococcus neoformans to ingestion by Acanthamoeba castellanii and macrophages provides insights into the evolutionary adaptation to the mammalian host
Abstract
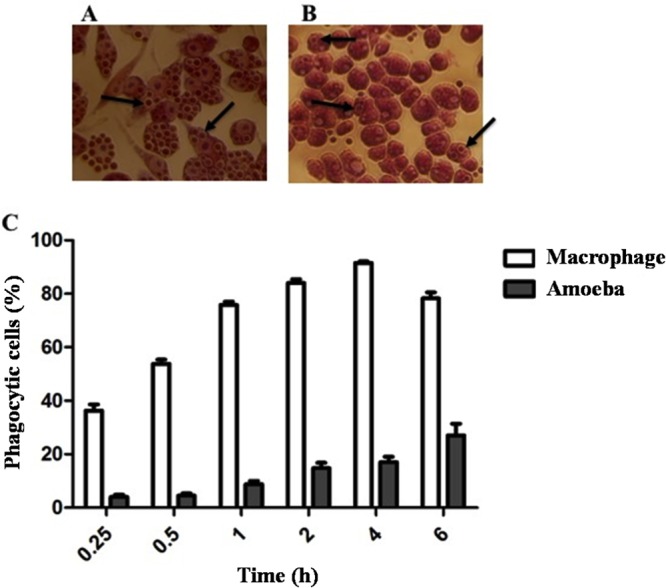
Virulence of Cryptococcus neoformans for mammals, and in particular its intracellular style, was proposed to emerge from evolutionary pressures on its natural environment by protozoan predation, which promoted the selection of strategies that allow intracellular survival in macrophages. In fact, Acanthamoeba castellanii ingests yeast cells, which then can replicate intracellularly. In addition, most fungal factors needed to establish infection in the mammalian host are also important for survival within the amoeba. To better understand the origin of C. neoformans virulence, we compared the transcriptional profile of yeast cells internalized by amoebae and murine macrophages after 6 h of infection. Our results showed 656 and 293 genes whose expression changed at least 2-fold in response to the intracellular environments of amoebae and macrophages, respectively. Among the genes that were found in both groups, we focused on open reading frame (ORF) CNAG_05662, which was potentially related to sugar transport but had no determined biological function. To characterize its function, we constructed a mutant strain and evaluated its ability to grow on various carbon sources. The results showed that this gene, named PTP1 (polyol transporter protein 1), is involved in the transport of 5- and 6-carbon polyols such as mannitol and sorbitol, but its presence or absence had no effect on cryptococcal virulence for mice or moth larvae. Overall, these results are consistent with the hypothesis that the capacity for mammalian virulence originated from fungus-protozoan interactions in the environment and provide a better understanding of how C. neoformans adapts to the mammalian host.
Figures






References
-
- Ellis DH, Pfeiffer TJ. 1990. Ecology, life cycle, and infectious propagule of Cryptococcus neoformans. Lancet 336:923–925 - PubMed
-
- Alanio A, Desnos-Ollivier M, Dromer F. 9 August 2011. Dynamics of Cryptococcus neoformans-macrophage interactions reveal that fungal background influences outcome during cryptococcal meningoencephalitis in humans. mBio 2(4):e00158-11 doi:10.1128/mBio.00158-11 - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
