

Chemosensory brush cells of the trachea. A stable population in a dynamic epithelium
- PMID: 23526223
- PMCID: PMC3824035
- DOI: 10.1165/rcmb.2012-0485OC
Chemosensory brush cells of the trachea. A stable population in a dynamic epithelium
Abstract
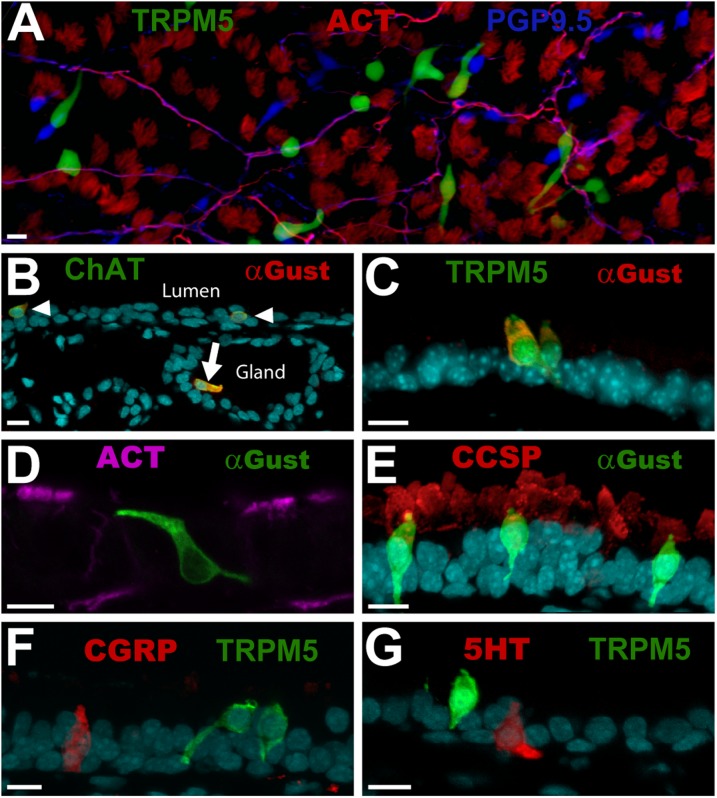
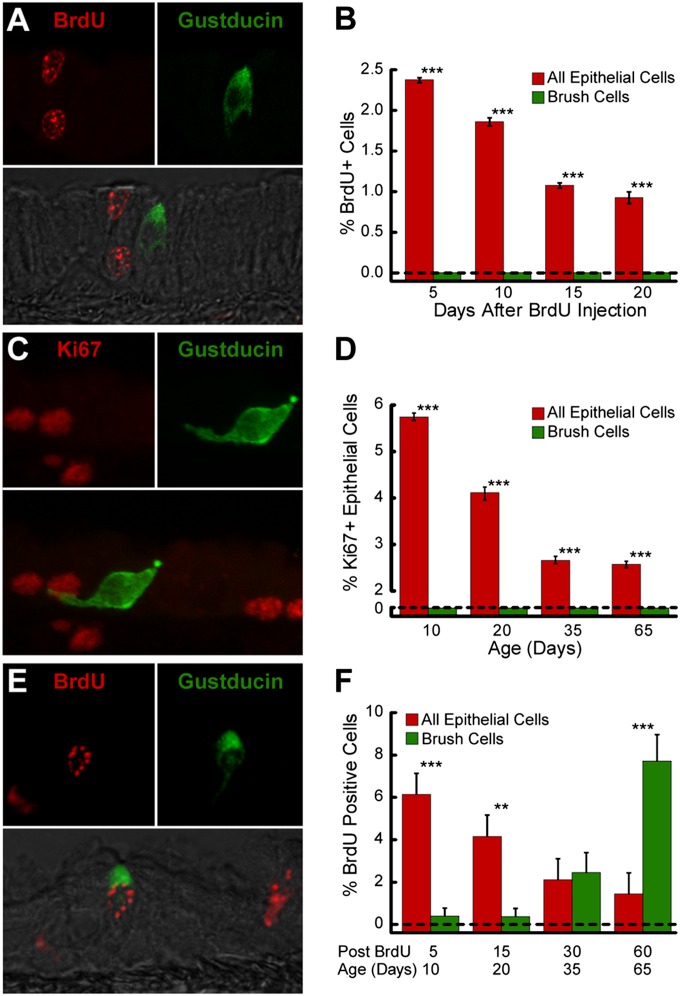
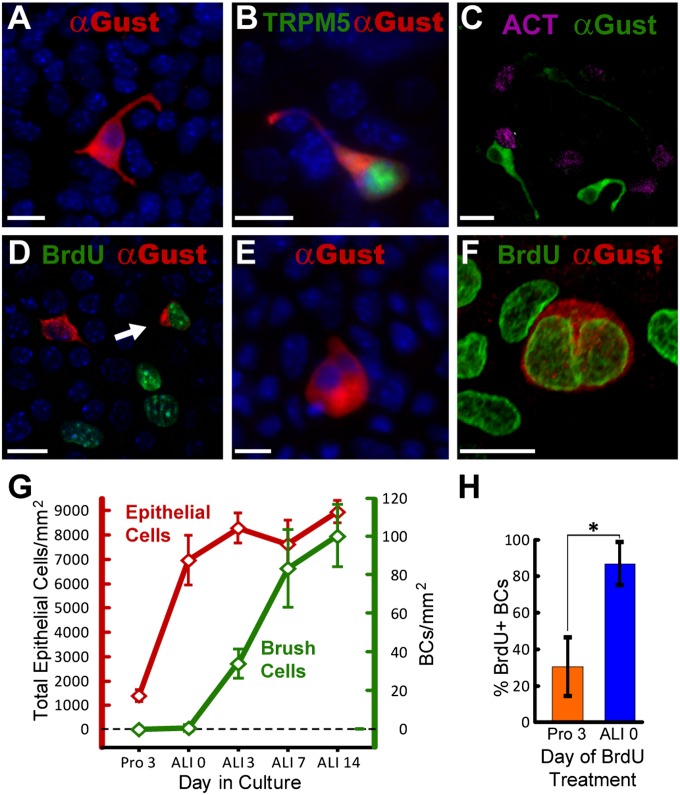
Tracheal brush cells (BCs) are specialized epithelial chemosensors that use the canonical taste transduction cascade to detect irritants. To test whether BCs are replaced at the same rate as other cells in the surrounding epithelium of adult mice, we used 5-bromo-2'-deoxyuridine (BrdU) to label dividing cells. Although scattered BrdU-labeled epithelial cells are present 5-20 days after BrdU, no BCs are labeled. These data indicate that BCs comprise a relatively static population. To determine how BCs are generated during development, we injected 5-day-old mice with BrdU and found labeled BCs and non-BC epithelial cells 5 days after BrdU. During the next 60 days, the percentage of labeled BCs increased, whereas the percentage of other labeled cell types decreased. These data suggest that BCs are generated from non-BC progenitor cells during postnatal tracheal growth. To test whether the adult epithelium retains the capacity to generate BCs, tracheal epithelial cells were recovered from adult mice and grown in an air-liquid interface (ALI) culture. After transition to differentiation conditions, BCs are detected, and comprise 1% of the total cell population by Day 14. BrdU added to cultures before the differentiation of BCs was chased into BCs, indicating that the increase in BC density is attributable to the proliferation of a non-BC progenitor. We conclude that: (1) BCs are normally a static population in adult mice; (2) BC progenitors proliferate and differentiate during neonatal development; and (3) BCs can be regenerated from a proliferative population resident in adult epithelium.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
