

Young lives lost as B cells falter: what we are learning about antibody responses in malaria
- PMID: 23526829
- PMCID: PMC3608210
- DOI: 10.4049/jimmunol.1203067
Young lives lost as B cells falter: what we are learning about antibody responses in malaria
Abstract
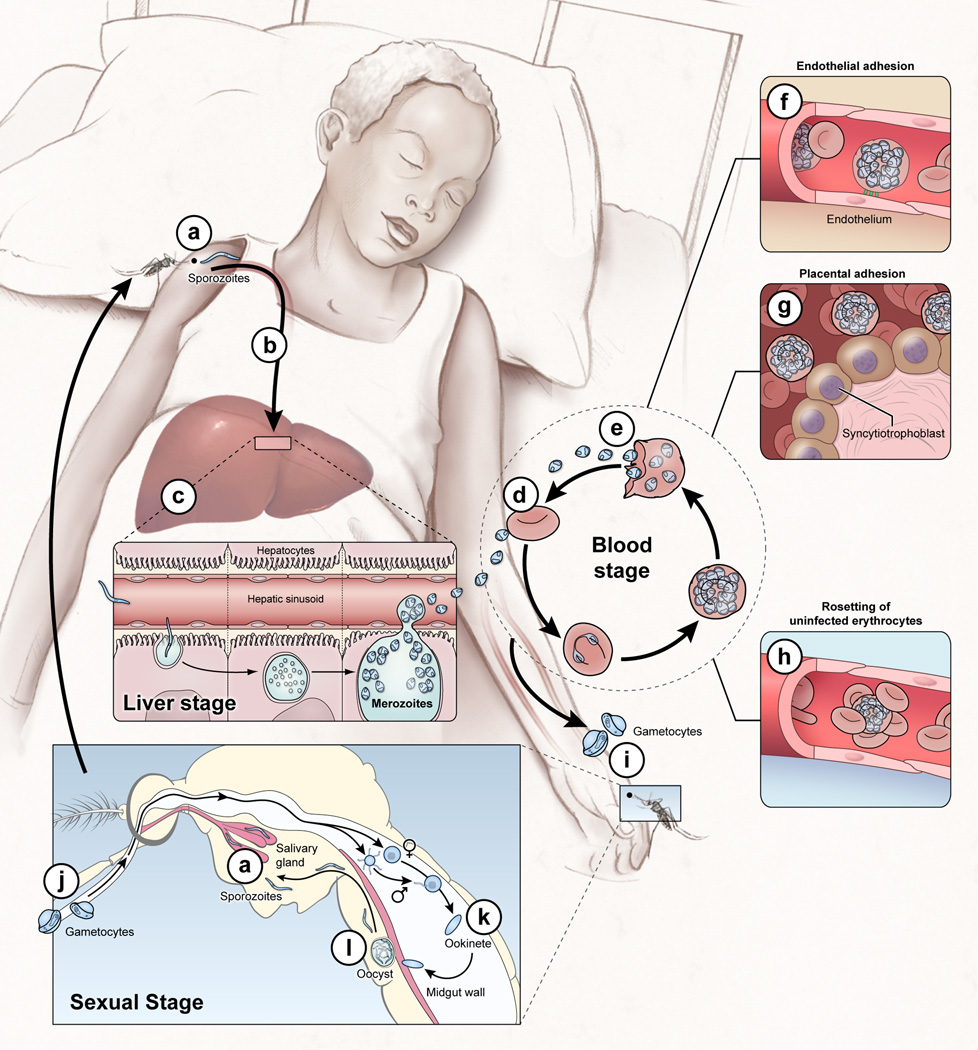
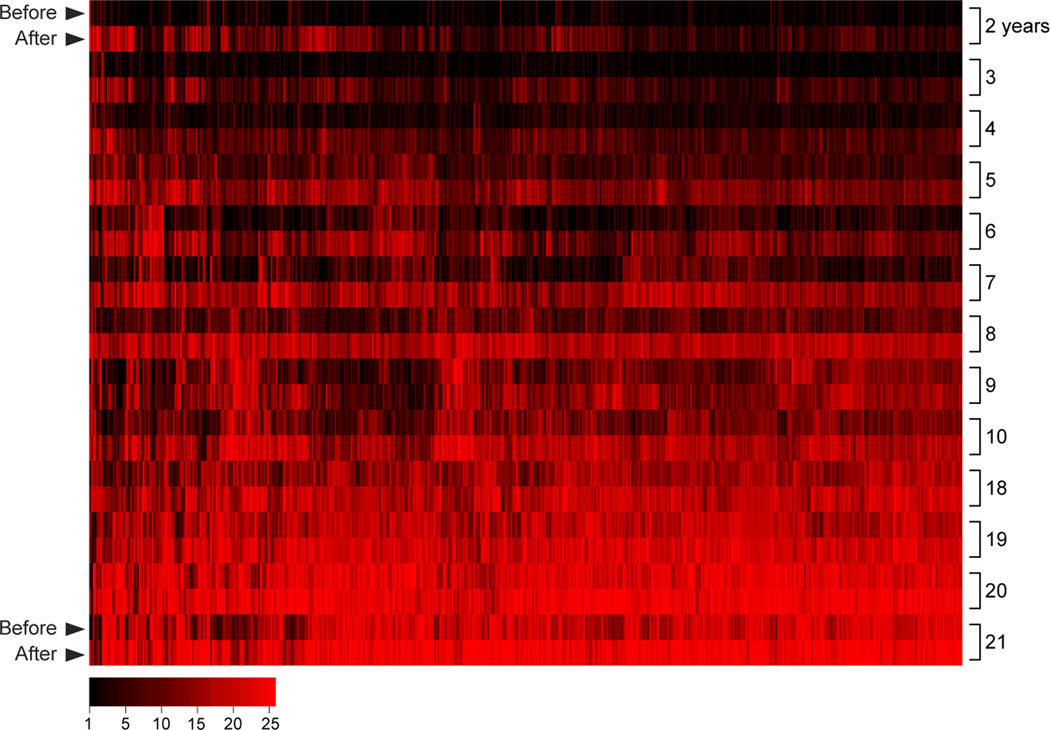
Plasmodium falciparum malaria remains a major public health threat for which there is no licensed vaccine. Abs play a key role in malaria immunity, but Ab-mediated protection is only acquired after years of repeated infections, leaving children in endemic areas vulnerable to severe malaria and death. Many P. falciparum Ags are extraordinarily diverse and clonally variant, which likely contribute to the inefficient acquisition of protective Abs. However, mounting evidence suggests that there is more to the story and that infection-induced dysregulation of B cell function also plays a role. We herein review progress toward understanding the B cell biology of P. falciparum infection, focusing on what has been learned from population-based studies in malaria-endemic areas. We suggest ways in which advances in immunology and genomics-based technology can further improve our understanding of the B cell response in malaria and perhaps illuminate new pathways to the development of effective vaccines.
Figures
References
-
- Murray CJ, Rosenfeld LC, Lim SS, Andrews KG, Foreman KJ, Haring D, Fullman N, Naghavi M, Lozano R, Lopez AD. Global malaria mortality between 1980 and 2010: a systematic analysis. Lancet. 2012;379:413–431. - PubMed
-
- Miller LH, Baruch DI, Marsh K, Doumbo OK. The pathogenic basis of malaria. Nature. 2002;415:673–679. - PubMed
-
- Marsh K, Kinyanjui S. Immune effector mechanisms in malaria. Parasite Immunol. 2006;28:51–60. - PubMed
-
- Jelinek T, Schulte C, Behrens R, Grobusch MP, Coulaud JP, Bisoffi Z, Matteelli A, Clerinx J, Corachan M, Puente S, Gjorup I, Harms G, Kollaritsch H, Kotlowski A, Bjorkmann A, Delmont JP, Knobloch J, Nielsen LN, Cuadros J, Hatz C, Beran J, Schmid ML, Schulze M, Lopez-Velez R, Fleischer K, Kapaun A, McWhinney P, Kern P, Atougia J, Fry G, da Cunha S, Boecken G. Imported Falciparum malaria in Europe: sentinel surveillance data from the European network on surveillance of imported infectious diseases. Clin Infect Dis. 2002;34:572–576. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
