

Galectin-3 functions as an alarmin: pathogenic role for sepsis development in murine respiratory tularemia
- PMID: 23527230
- PMCID: PMC3603908
- DOI: 10.1371/journal.pone.0059616
Galectin-3 functions as an alarmin: pathogenic role for sepsis development in murine respiratory tularemia
Abstract
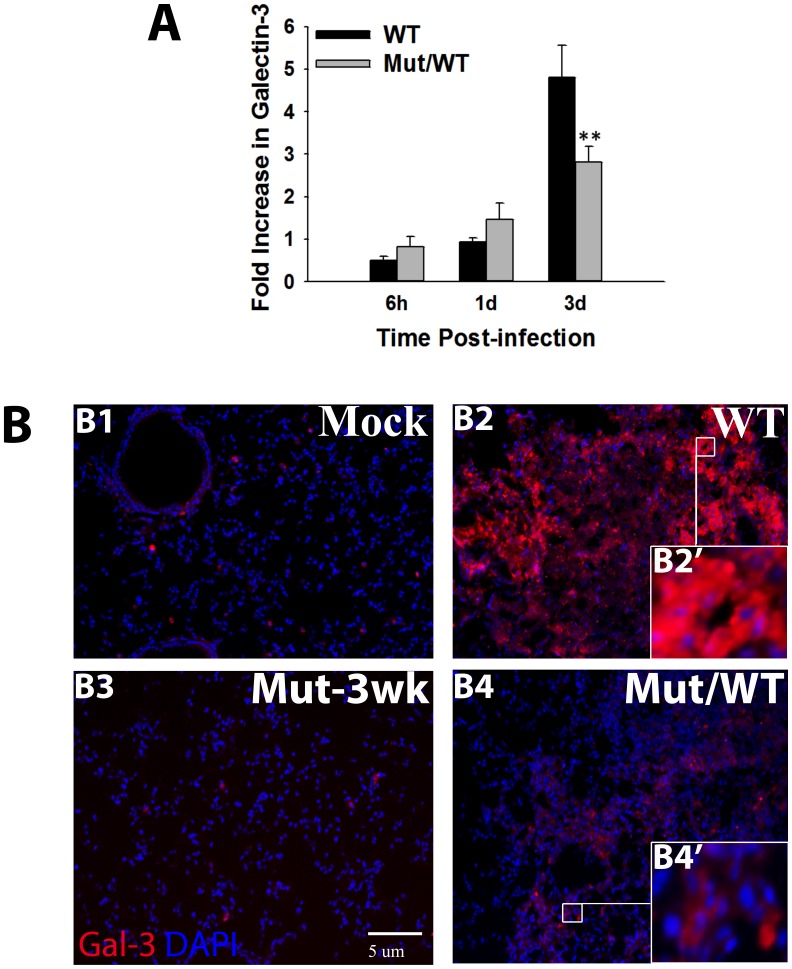
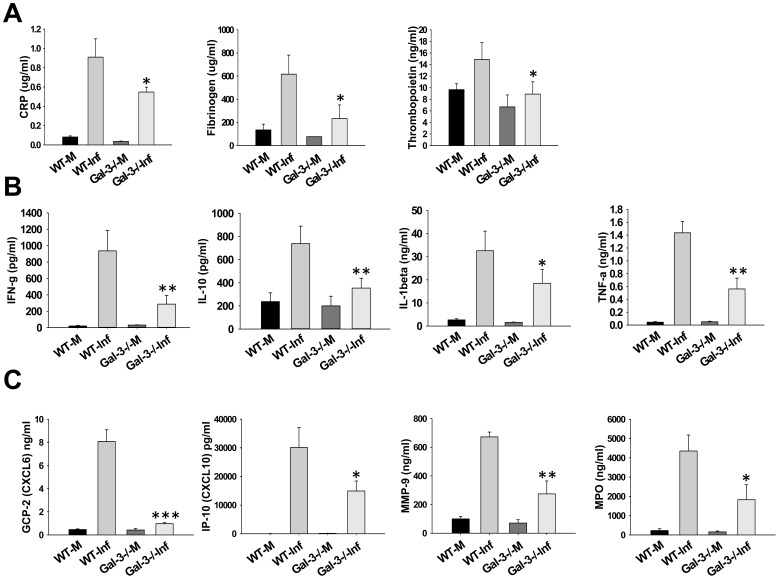
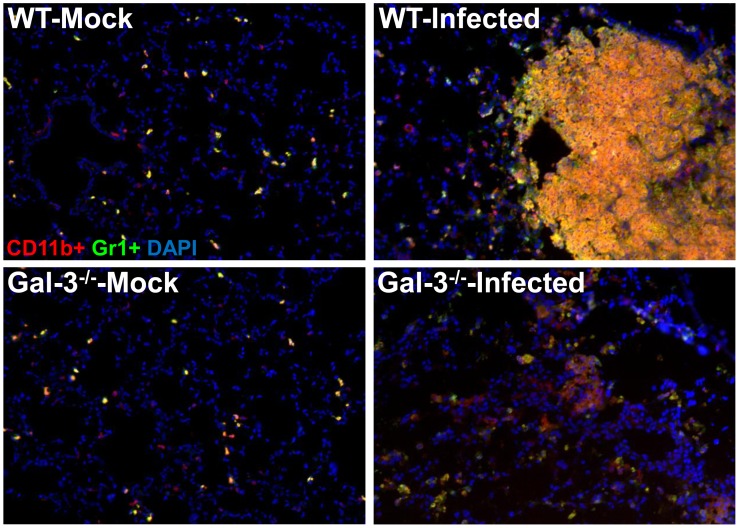
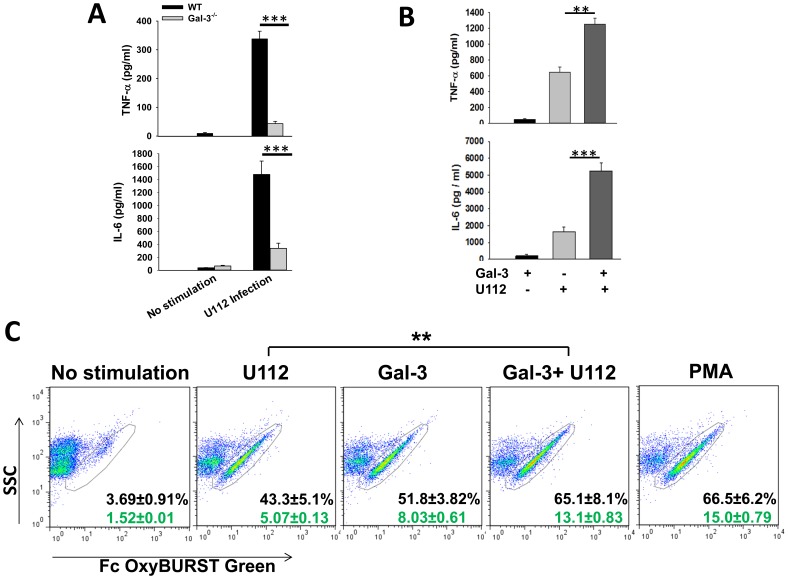
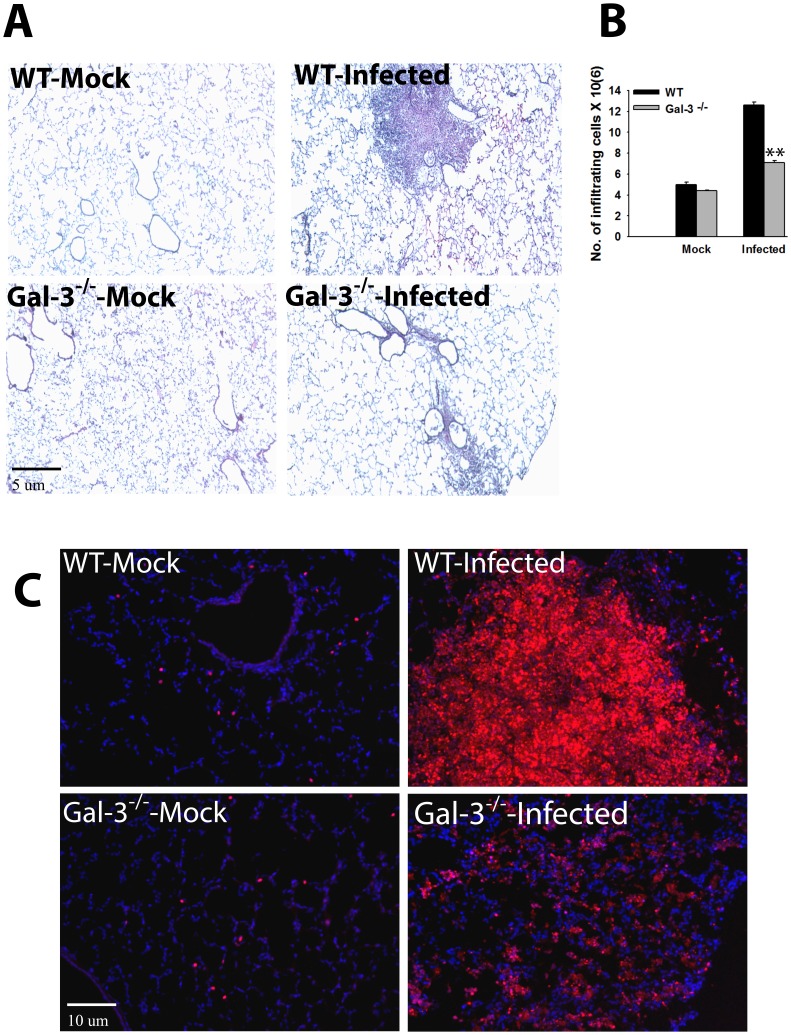
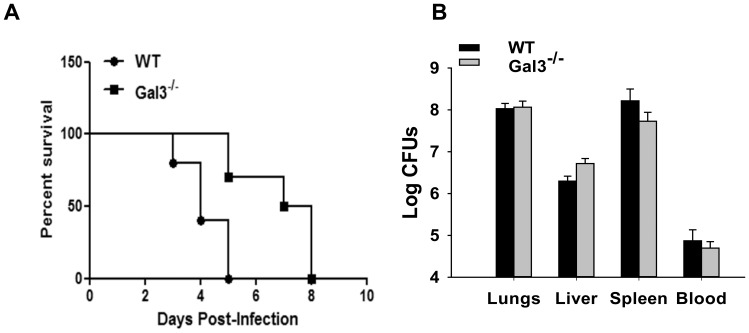
Sepsis is a complex immune disorder with a mortality rate of 20-50% and currently has no therapeutic interventions. It is thus critical to identify and characterize molecules/factors responsible for its development. We have recently shown that pulmonary infection with Francisella results in sepsis development. As extensive cell death is a prominent feature of sepsis, we hypothesized that host endogenous molecules called alarmins released from dead or dying host cells cause a hyperinflammatory response culminating in sepsis development. In the current study we investigated the role of galectin-3, a mammalian β-galactoside binding lectin, as an alarmin in sepsis development during F. novicida infection. We observed an upregulated expression and extracellular release of galectin-3 in the lungs of mice undergoing lethal pulmonary infection with virulent strain of F. novicida but not in those infected with a non-lethal, attenuated strain of the bacteria. In comparison with their wild-type C57Bl/6 counterparts, F. novicida infected galectin-3 deficient (galectin-3(-/-)) mice demonstrated significantly reduced leukocyte infiltration, particularly neutrophils in their lungs. They also exhibited a marked decrease in inflammatory cytokines, vascular injury markers, and neutrophil-associated inflammatory mediators. Concomitantly, in-vitro pre-treatment of primary neutrophils and macrophages with recombinant galectin-3 augmented F. novicida-induced activation of these cells. Correlating with the reduced inflammatory response, F. novicida infected galectin-3(-/-) mice exhibited improved lung architecture with reduced cell death and improved survival over wild-type mice, despite similar bacterial burden. Collectively, these findings suggest that galectin-3 functions as an alarmin by augmenting the inflammatory response in sepsis development during pulmonary F. novicida infection.
Conflict of interest statement
Figures
References
-
- Hotchkiss RS, Karl IE (2003) The pathophysiology and treatment of sepsis. N Engl J Med 348: 138–150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
