

Membrane fission reactions of the mammalian ESCRT pathway
- PMID: 23527693
- PMCID: PMC4047973
- DOI: 10.1146/annurev-biochem-072909-101058
Membrane fission reactions of the mammalian ESCRT pathway
Abstract
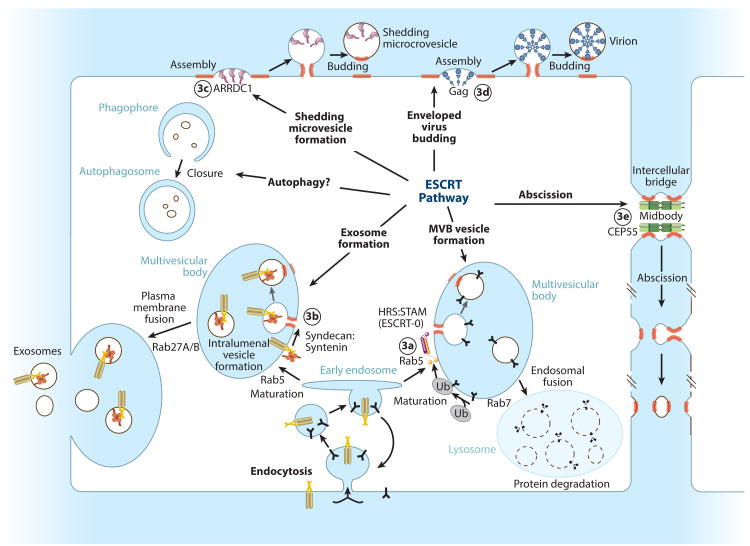
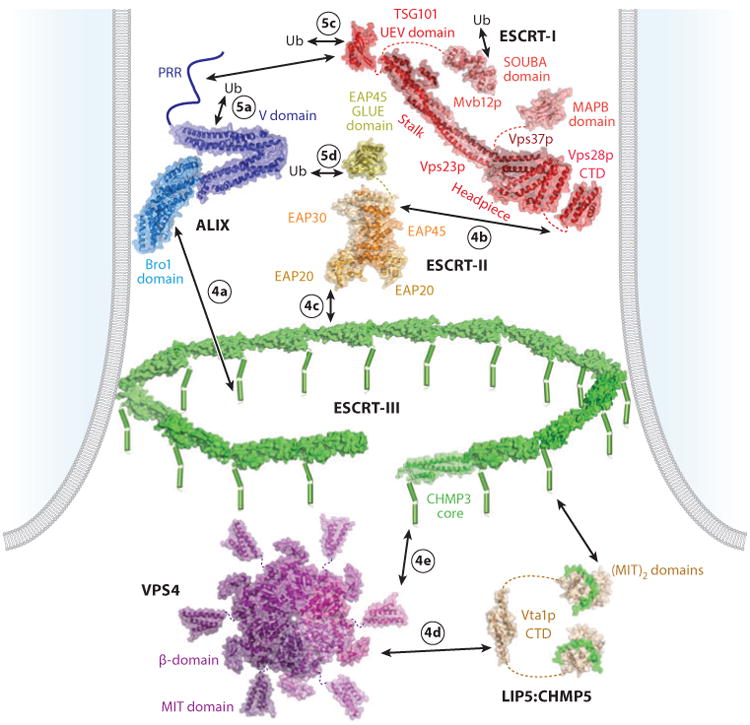
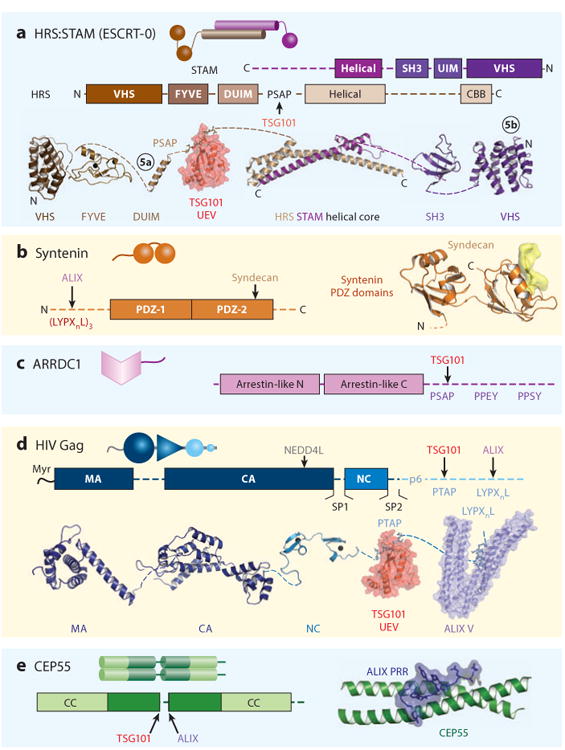
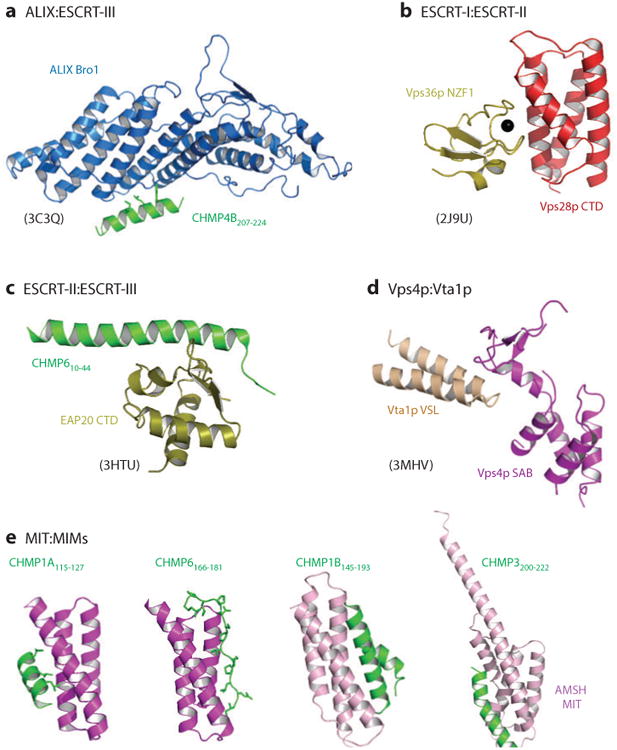
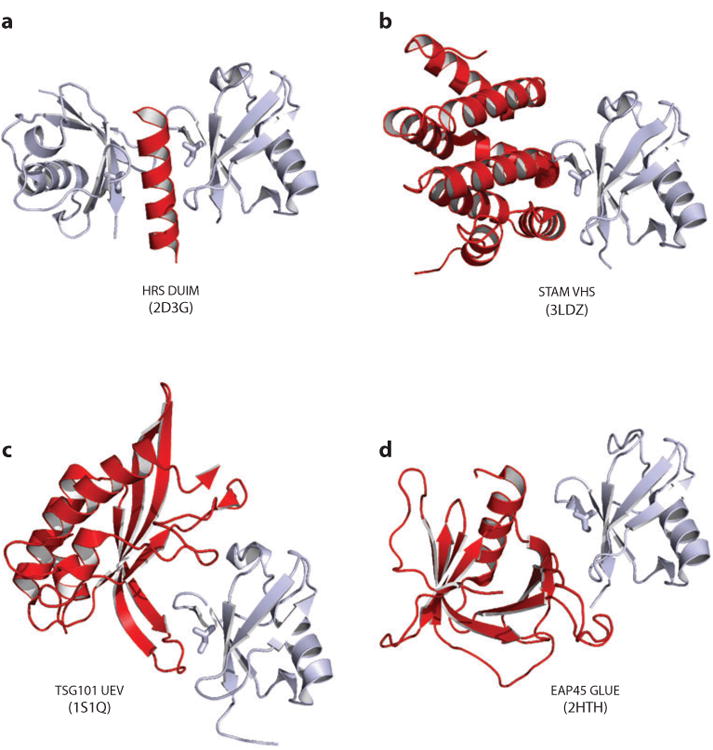
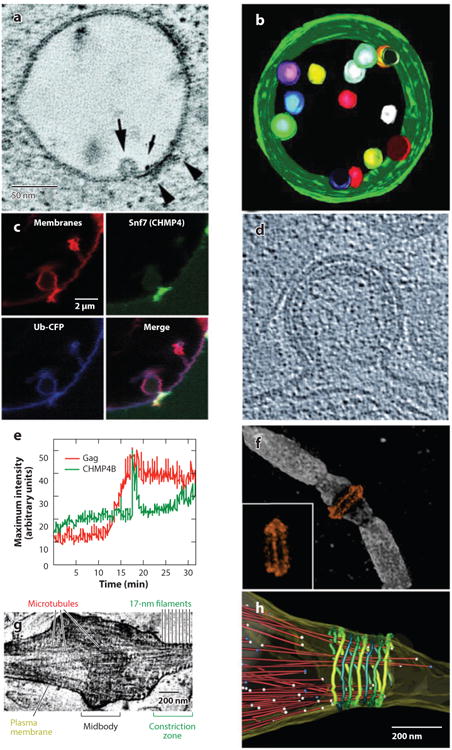
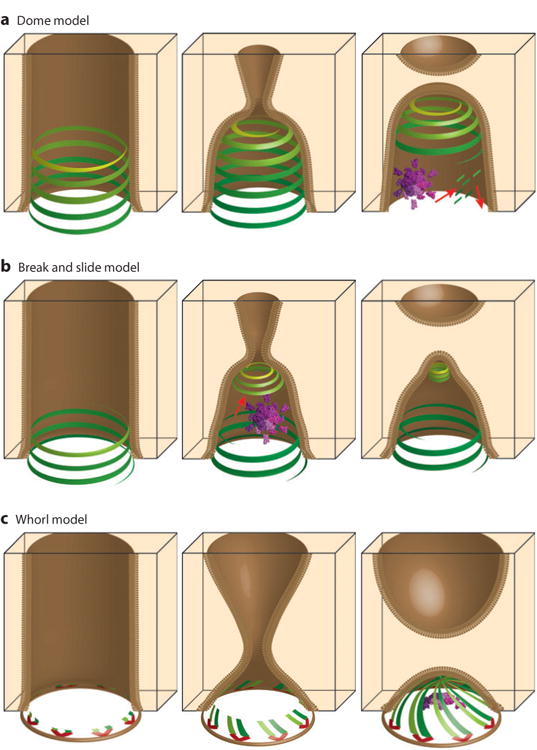
The endosomal sorting complexes required for transport (ESCRT) pathway was initially defined in yeast genetic screens that identified the factors necessary to sort membrane proteins into intraluminal endosomal vesicles. Subsequent studies have revealed that the mammalian ESCRT pathway also functions in a series of other key cellular processes, including formation of extracellular microvesicles, enveloped virus budding, and the abscission stage of cytokinesis. The core ESCRT machinery comprises Bro1 family proteins and ESCRT-I, ESCRT-II, ESCRT-III, and VPS4 complexes. Site-specific adaptors recruit these soluble factors to assemble on different cellular membranes, where they carry out membrane fission reactions. ESCRT-III proteins form filaments that draw membranes together from the cytoplasmic face, and mechanistic models have been advanced to explain how ESCRT-III filaments and the VPS4 ATPase can work together to catalyze membrane fission.
Figures
References
-
- Henne WM, Buchkovich NJ, Emr SD. The ESCRT pathway. Dev Cell. 2011;21:77–91. - PubMed
-
- Raiborg C, Stenmark H. Hrs and endocytic sorting of ubiquitinated membrane proteins. Cell Struct Funct. 2002;27:403–8. - PubMed
-
- Clague MJ, Urbé S. Hrs function: Viruses provide the clue. Trends Cell Biol. 2003;13:603–6. - PubMed
-
- Baietti MF, Zhang Z, Mortier E, Melchior A, Degeest G, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol. 2012;14:677–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
