

Comparative proteomic analysis of normal and collagen IX null mouse cartilage reveals altered extracellular matrix composition and novel components of the collagen IX interactome
- PMID: 23530037
- PMCID: PMC3650385
- DOI: 10.1074/jbc.M112.444810
Comparative proteomic analysis of normal and collagen IX null mouse cartilage reveals altered extracellular matrix composition and novel components of the collagen IX interactome
Abstract
Background: Collagen IX is an integral cartilage extracellular matrix component important in skeletal development and joint function.
Results: Proteomic analysis and validation studies revealed novel alterations in collagen IX null cartilage.
Conclusion: Matrilin-4, collagen XII, thrombospondin-4, fibronectin, βig-h3, and epiphycan are components of the in vivo collagen IX interactome.
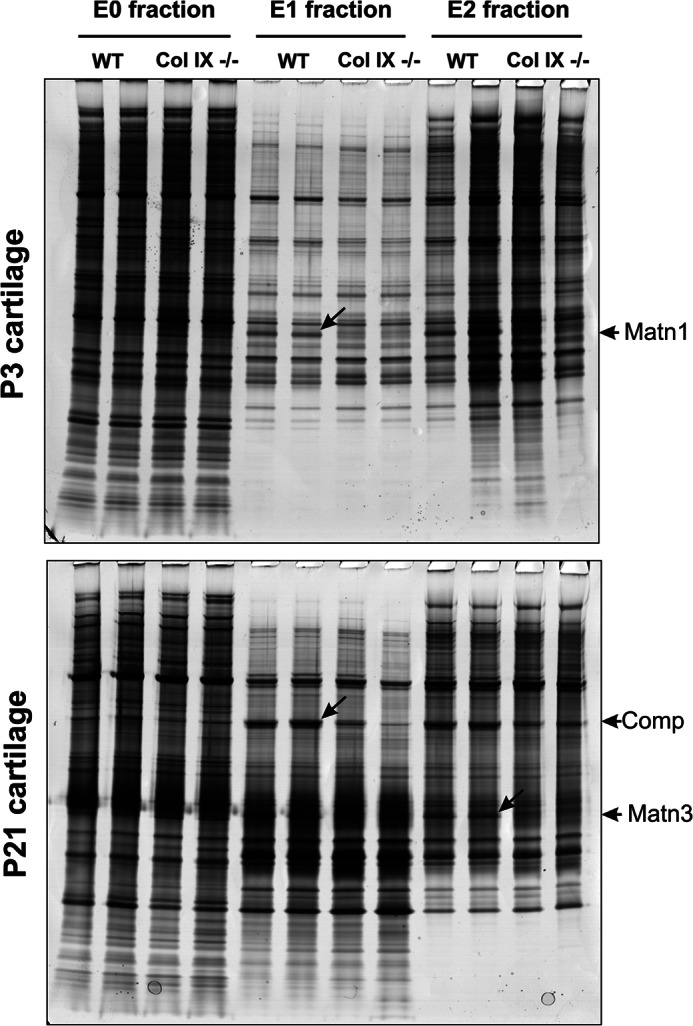
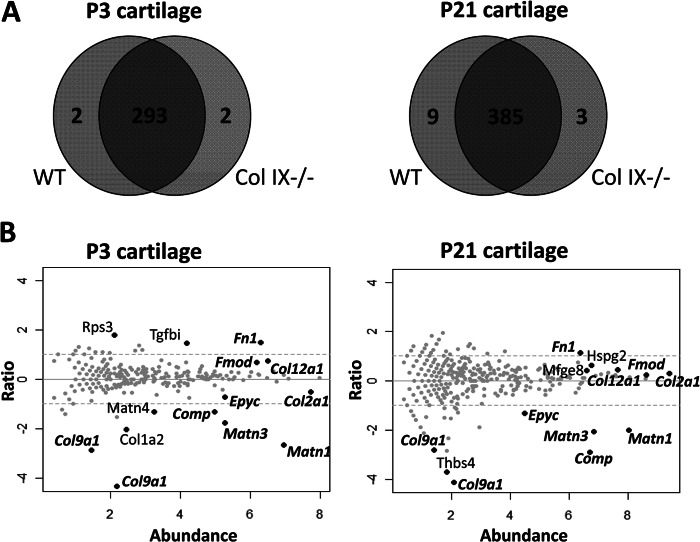
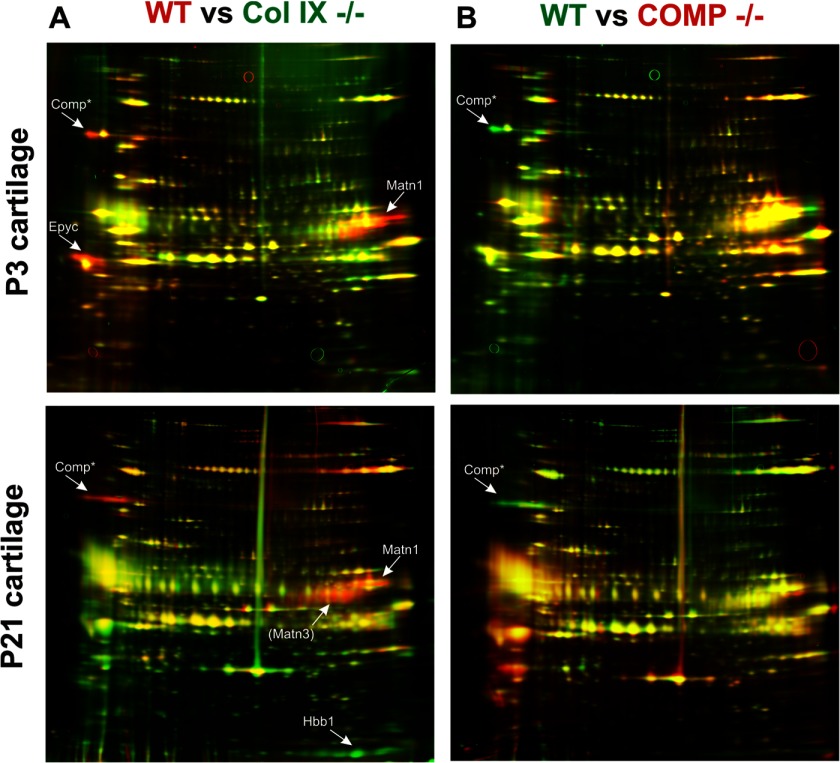
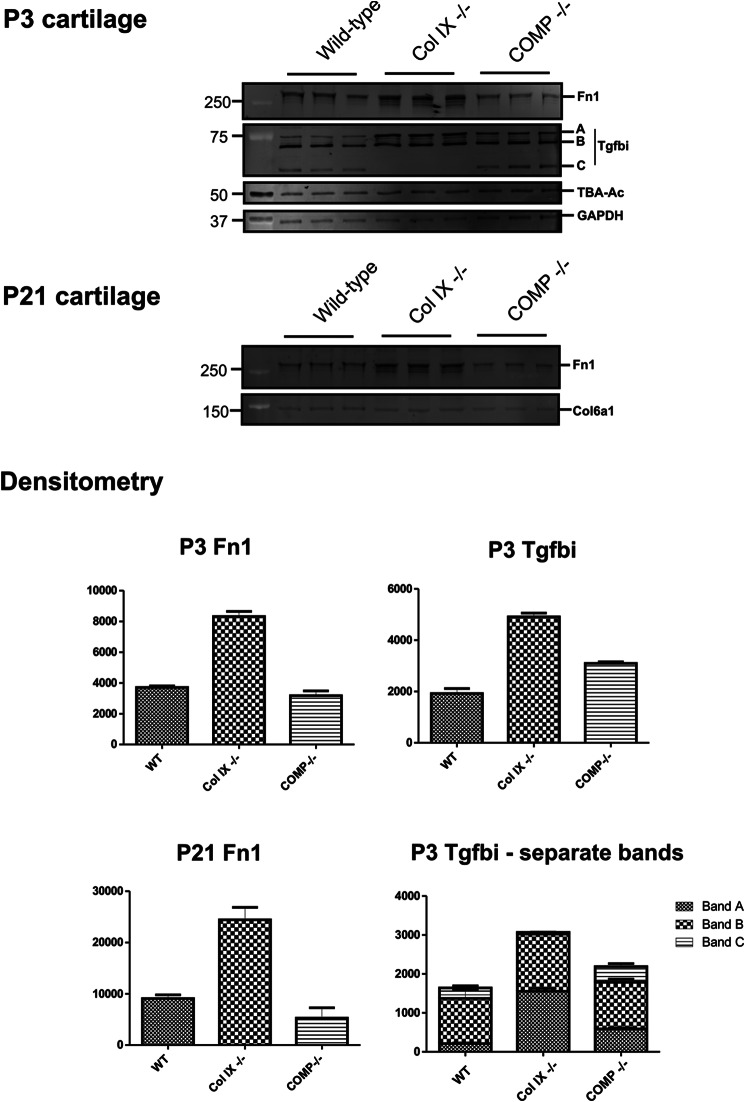
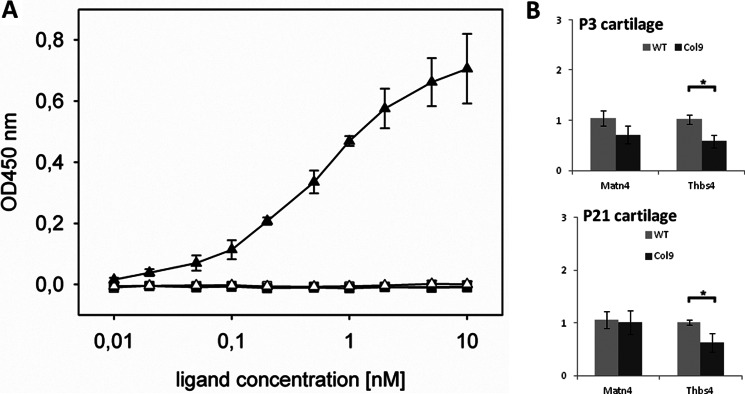
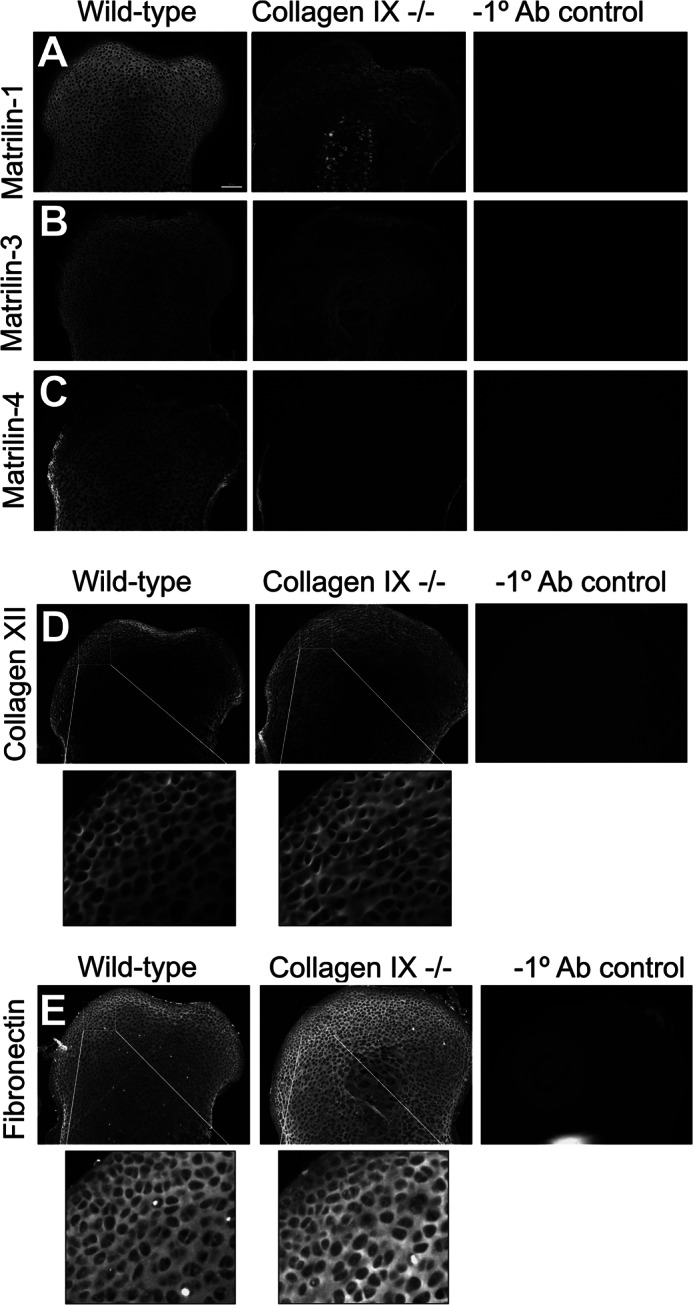
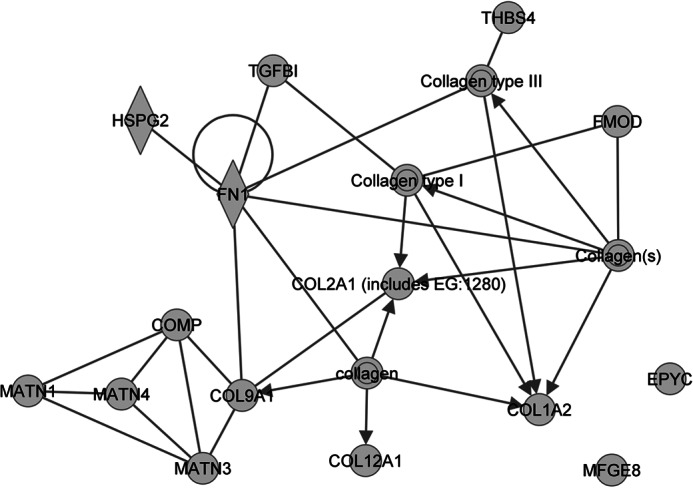
Significance: We applied a proteomics approach to advance our understanding of collagen IX ablation in cartilage. The cartilage extracellular matrix is essential for endochondral bone development and joint function. In addition to the major aggrecan/collagen II framework, the interacting complex of collagen IX, matrilin-3, and cartilage oligomeric matrix protein (COMP) is essential for cartilage matrix stability, as mutations in Col9a1, Col9a2, Col9a3, Comp, and Matn3 genes cause multiple epiphyseal dysplasia, in which patients develop early onset osteoarthritis. In mice, collagen IX ablation results in severely disturbed growth plate organization, hypocellular regions, and abnormal chondrocyte shape. This abnormal differentiation is likely to involve altered cell-matrix interactions but the mechanism is not known. To investigate the molecular basis of the collagen IX null phenotype we analyzed global differences in protein abundance between wild-type and knock-out femoral head cartilage by capillary HPLC tandem mass spectrometry. We identified 297 proteins in 3-day cartilage and 397 proteins in 21-day cartilage. Components that were differentially abundant between wild-type and collagen IX-deficient cartilage included 15 extracellular matrix proteins. Collagen IX ablation was associated with dramatically reduced COMP and matrilin-3, consistent with known interactions. Matrilin-1, matrilin-4, epiphycan, and thrombospondin-4 levels were reduced in collagen IX null cartilage, providing the first in vivo evidence for these proteins belonging to the collagen IX interactome. Thrombospondin-4 expression was reduced at the mRNA level, whereas matrilin-4 was verified as a novel collagen IX-binding protein. Furthermore, changes in TGFβ-induced protein βig-h3 and fibronectin abundance were found in the collagen IX knock-out but not associated with COMP ablation, indicating specific involvement in the abnormal collagen IX null cartilage. In addition, the more widespread expression of collagen XII in the collagen IX-deficient cartilage suggests an attempted compensatory response to the absence of collagen IX. Our differential proteomic analysis of cartilage is a novel approach to identify candidate matrix protein interactions in vivo, underpinning further analysis of mutant cartilage lacking other matrix components or harboring disease-causing mutations.
Keywords: Cartilage; Chondrocytes; Chondrodysplasia; Collagen; Extracellular Matrix; Mass Spectrometry (MS); Proteomics.
Figures
References
-
- Newman B., Wallis G. A. (2003) Skeletal dysplasias caused by a disruption of skeletal patterning and endochondral ossification. Clin. Genet. 63, 241–251 - PubMed
-
- Warman M. L., Cormier-Daire V., Hall C., Krakow D., Lachman R., LeMerrer M., Mortier G., Mundlos S., Nishimura G., Rimoin D. L., Robertson S., Savarirayan R., Sillence D., Spranger J., Unger S., Zabel B., Superti-Furga A. (2011) Nosology and classification of genetic skeletal disorders. 2010 revision. Am. J. Med. Genet. A. 155A, 943–968 - PMC - PubMed
-
- Eyre D. R., Weis M. A., Wu J. J. (2006) Articular cartilage collagen. An irreplaceable framework? Eur. Cell. Mater. 12, 57–63 - PubMed
-
- Briggs M. D., Chapman K. L. (2002) Pseudoachondroplasia and multiple epiphyseal dysplasia. Mutation review, molecular interactions, and genotype to phenotype correlations. Hum. Mutat. 19, 465–478 - PubMed
-
- Bönnemann C. G., Cox G. F., Shapiro F., Wu J. J., Feener C. A., Thompson T. G., Anthony D. C., Eyre D. R., Darras B. T., Kunkel L. M. (2000) A mutation in the α3 chain of type IX collagen causes autosomal dominant multiple epiphyseal dysplasia with mild myopathy. Proc. Natl. Acad. Sci. U.S.A. 97, 1212–1217 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
