

MCJ/DnaJC15, an endogenous mitochondrial repressor of the respiratory chain that controls metabolic alterations
- PMID: 23530063
- PMCID: PMC3648061
- DOI: 10.1128/MCB.00189-13
MCJ/DnaJC15, an endogenous mitochondrial repressor of the respiratory chain that controls metabolic alterations
Abstract
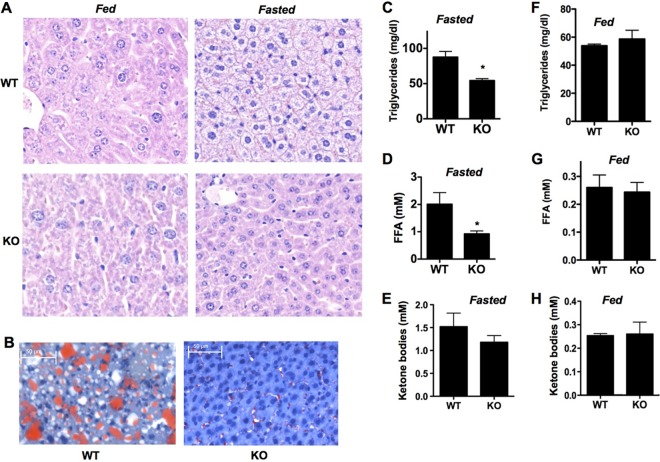
Mitochondria are the main engine that generates ATP through oxidative phosphorylation within the respiratory chain. Mitochondrial respiration is regulated according to the metabolic needs of cells and can be modulated in response to metabolic changes. Little is known about the mechanisms that regulate this process. Here, we identify MCJ/DnaJC15 as a distinct cochaperone that localizes at the mitochondrial inner membrane, where it interacts preferentially with complex I of the electron transfer chain. We show that MCJ impairs the formation of supercomplexes and functions as a negative regulator of the respiratory chain. The loss of MCJ leads to increased complex I activity, mitochondrial membrane potential, and ATP production. Although MCJ is dispensable for mitochondrial function under normal physiological conditions, MCJ deficiency affects the pathophysiology resulting from metabolic alterations. Thus, enhanced mitochondrial respiration in the absence of MCJ prevents the pathological accumulation of lipids in the liver in response to both fasting and a high-cholesterol diet. Impaired expression or loss of MCJ expression may therefore result in a "rapid" metabolism that mitigates the consequences of metabolic disorders.
Figures







References
-
- Cruciat CM, Brunner S, Baumann F, Neupert W, Stuart RA. 2000. The cytochrome bc1 and cytochrome c oxidase complexes associate to form a single supracomplex in yeast mitochondria. J. Biol. Chem. 275:18093–18098 - PubMed
-
- Stuart RA. 2008. Supercomplex organization of the oxidative phosphorylation enzymes in yeast mitochondria. J. Bioenerg. Biomembr. 40:411–417 - PubMed
-
- Acin-Perez R, Fernandez-Silva P, Peleato ML, Perez-Martos A, Enriquez JA. 2008. Respiratory active mitochondrial supercomplexes. Mol. Cell 32:529–539 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases