

Construction and validation of a homology model of the human voltage-gated proton channel hHV1
- PMID: 23530137
- PMCID: PMC3607825
- DOI: 10.1085/jgp.201210856
Construction and validation of a homology model of the human voltage-gated proton channel hHV1
Abstract
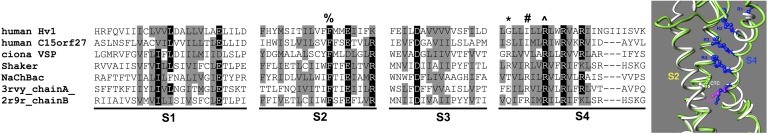
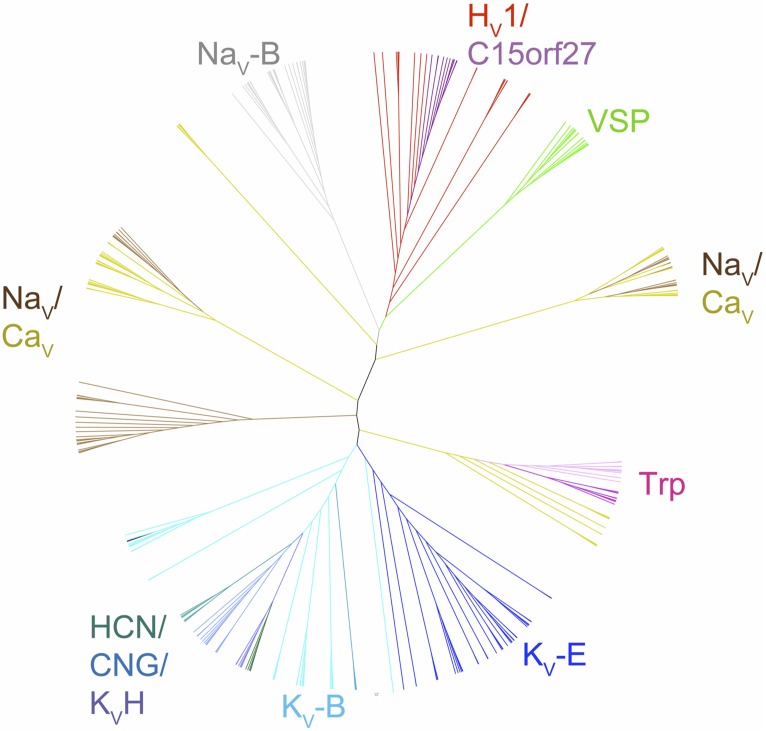
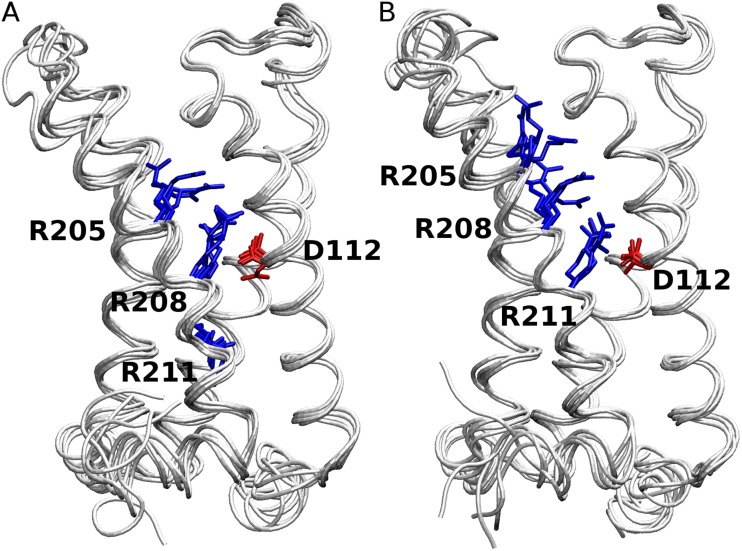
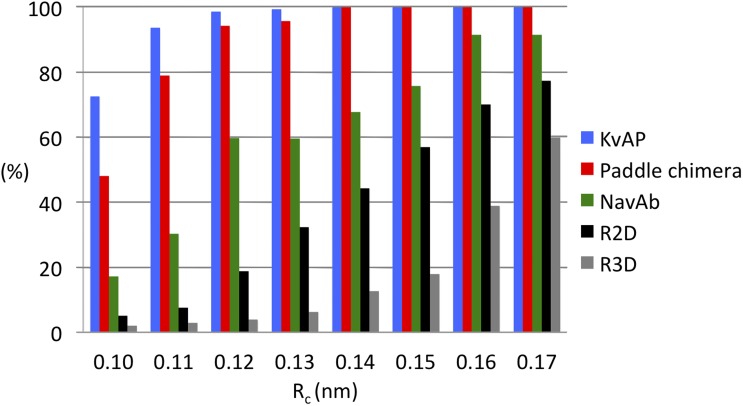
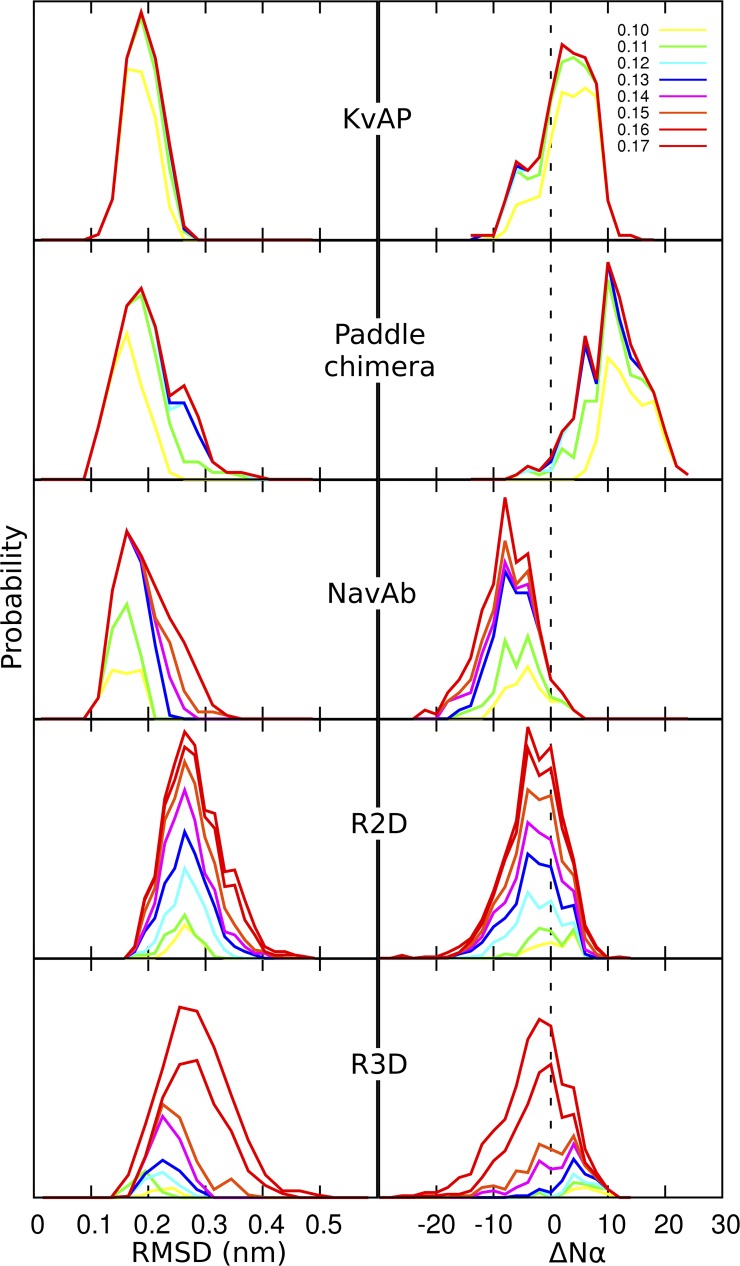
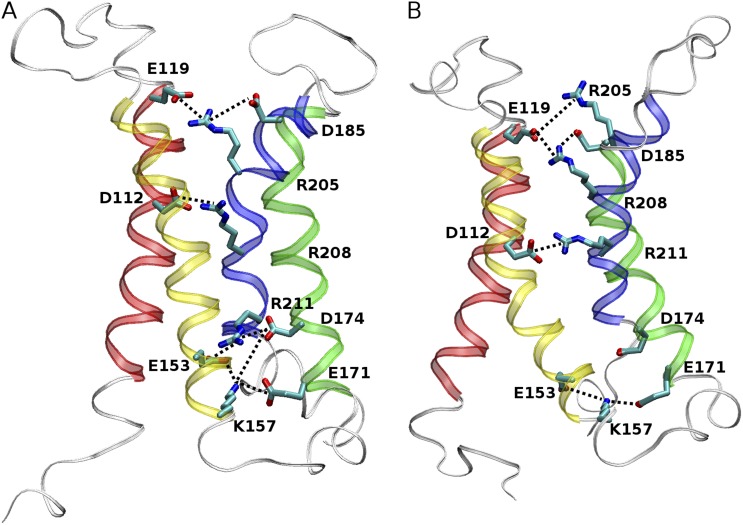
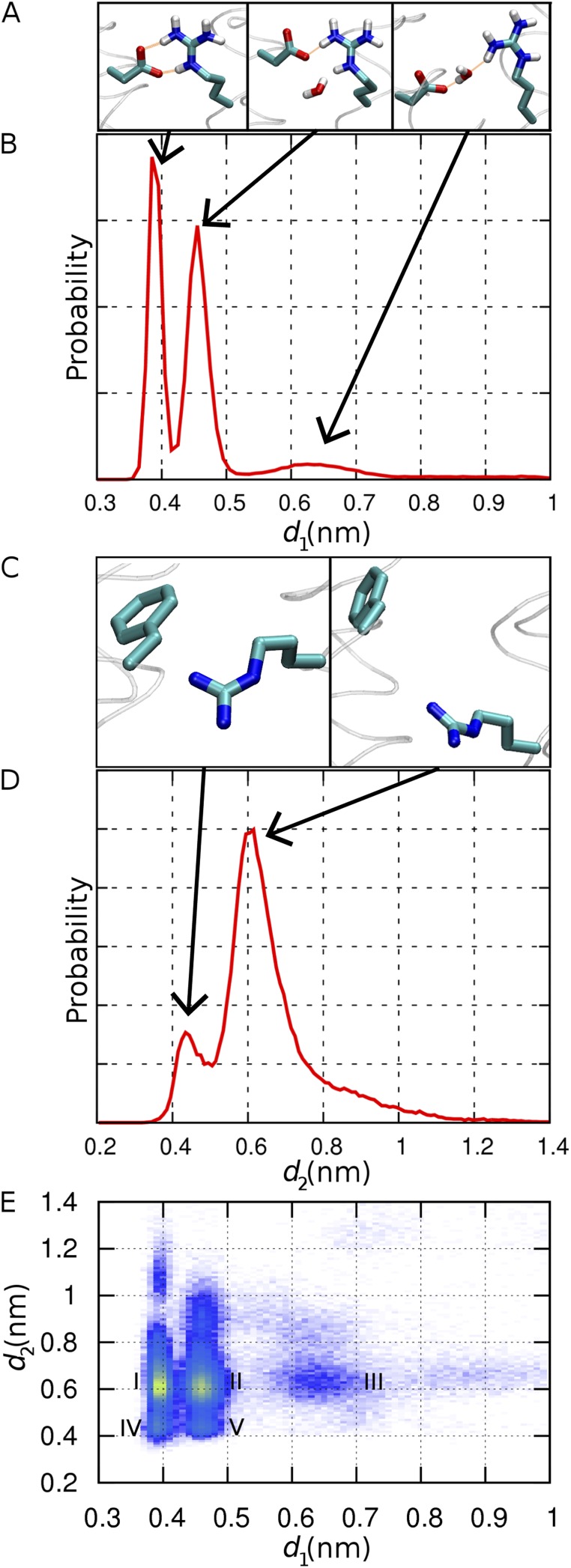
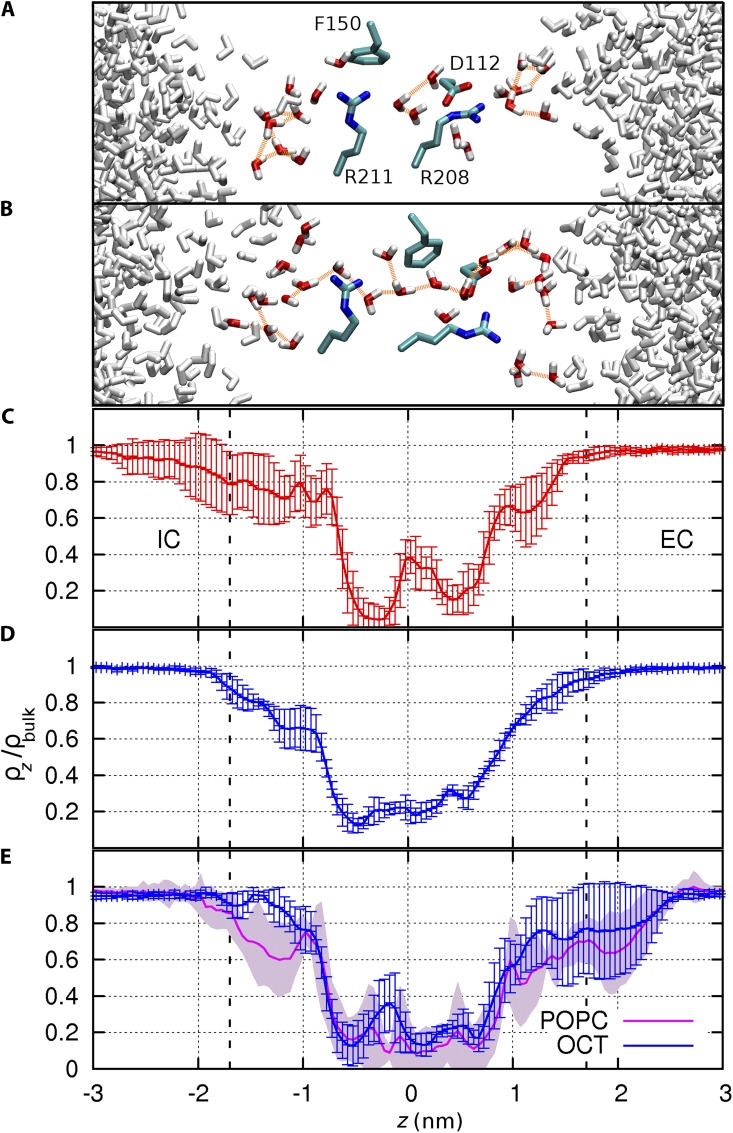
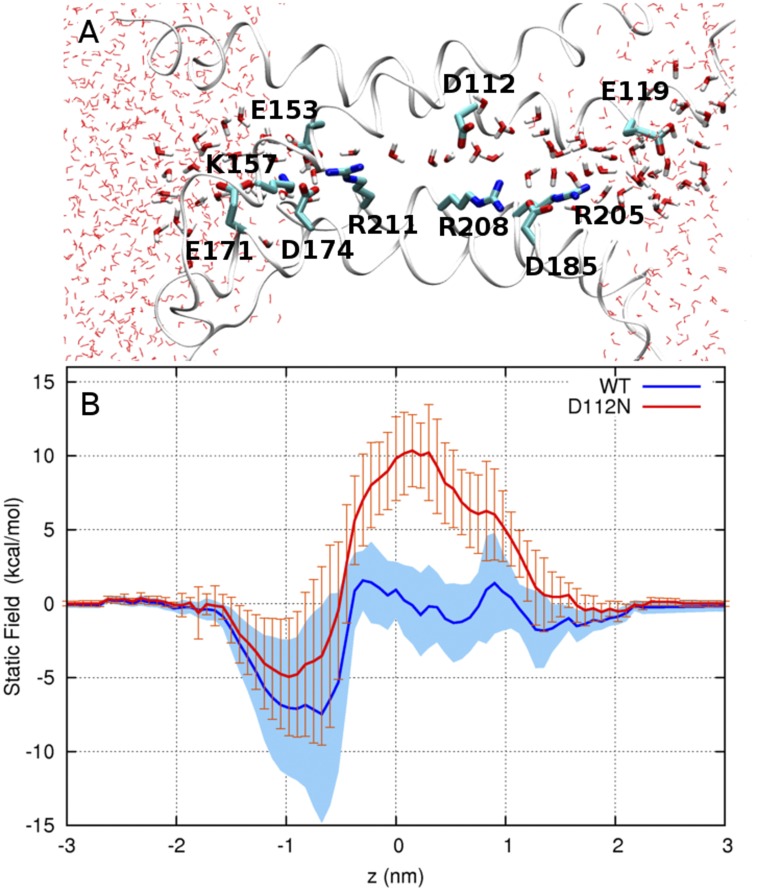
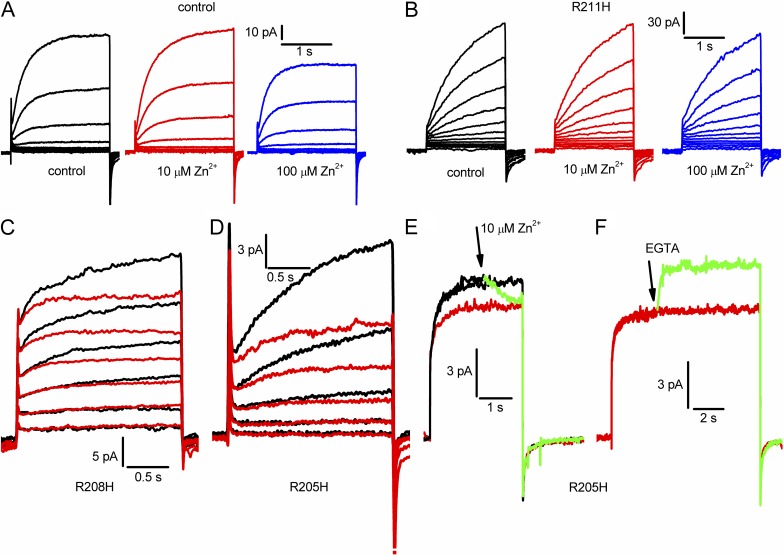
The topological similarity of voltage-gated proton channels (H(V)1s) to the voltage-sensing domain (VSD) of other voltage-gated ion channels raises the central question of whether H(V)1s have a similar structure. We present the construction and validation of a homology model of the human H(V)1 (hH(V)1). Multiple structural alignment was used to construct structural models of the open (proton-conducting) state of hH(V)1 by exploiting the homology of hH(V)1 with VSDs of K(+) and Na(+) channels of known three-dimensional structure. The comparative assessment of structural stability of the homology models and their VSD templates was performed using massively repeated molecular dynamics simulations in which the proteins were allowed to relax from their initial conformation in an explicit membrane mimetic. The analysis of structural deviations from the initial conformation based on up to 125 repeats of 100-ns simulations for each system reveals structural features consistently retained in the homology models and leads to a consensus structural model for hH(V)1 in which well-defined external and internal salt-bridge networks stabilize the open state. The structural and electrostatic properties of this open-state model are compatible with proton translocation and offer an explanation for the reversal of charge selectivity in neutral mutants of Asp(112). Furthermore, these structural properties are consistent with experimental accessibility data, providing a valuable basis for further structural and functional studies of hH(V)1. Each Arg residue in the S4 helix of hH(V)1 was replaced by His to test accessibility using Zn(2+) as a probe. The two outermost Arg residues in S4 were accessible to external solution, whereas the innermost one was accessible only to the internal solution. Both modeling and experimental data indicate that in the open state, Arg(211), the third Arg residue in the S4 helix in hH(V)1, remains accessible to the internal solution and is located near the charge transfer center, Phe(150).
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
