

Characterization of platelet aminophospholipid externalization reveals fatty acids as molecular determinants that regulate coagulation
- PMID: 23530199
- PMCID: PMC3625294
- DOI: 10.1073/pnas.1222419110
Characterization of platelet aminophospholipid externalization reveals fatty acids as molecular determinants that regulate coagulation
Abstract
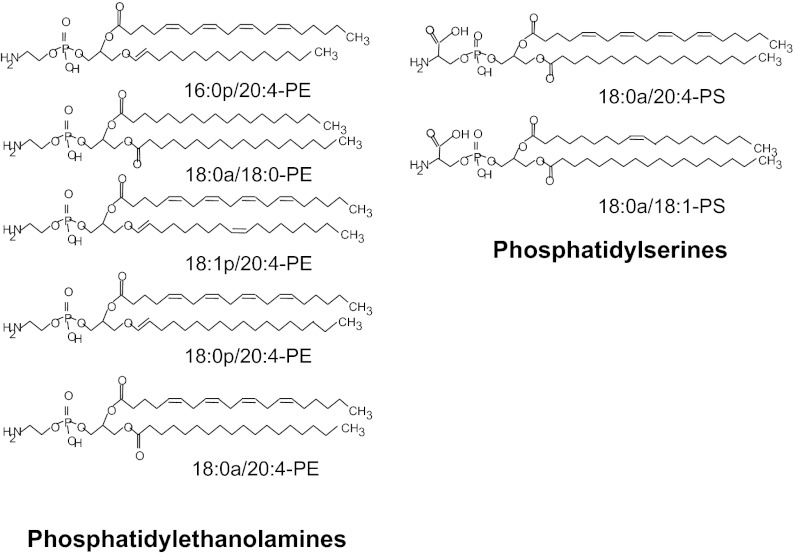
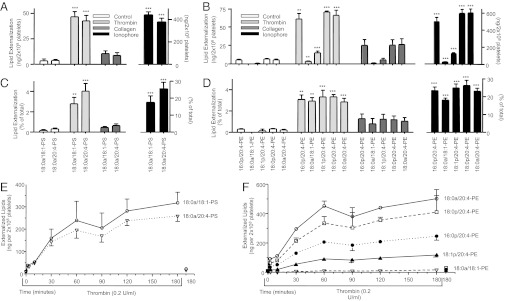
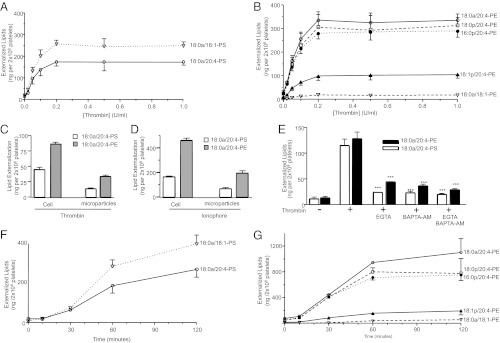
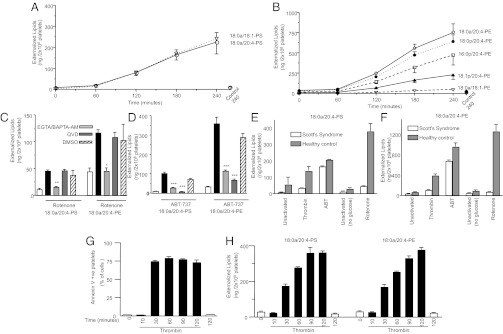
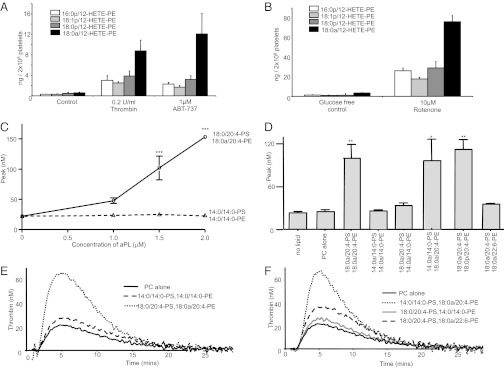
Aminophospholipid (APL) trafficking across the plasma membrane is a key event in cell activation, apoptosis, and aging and is required for clearance of dying cells and coagulation. Currently the phospholipid molecular species externalized are unknown. Using a lipidomic method, we show that thrombin, collagen, or ionophore-activated human platelets externalize two phosphatidylserines (PSs) and five phosphatidylethanolamines (PEs). Four percent of the total cellular PE/PS pool (∼300 ng/2 × 10(8) cells, thrombin), is externalized via calcium mobilization and protease-activated receptors-1 and -4, and 48% is contained in microparticles. Apoptosis and energy depletion (aging) externalized the same APLs in a calcium-dependent manner, and all stimuli externalized oxidized phospholipids, termed hydroxyeicosatetraenoic acid-PEs. Transmembrane protein-16F (TMEM-16F), the protein mutated in Scott syndrome, was required for PE/PS externalization during thrombin activation and energy depletion, but not apoptosis. Platelet-specific APLs optimally supported tissue factor-dependent coagulation in human plasma, vs. APL with longer or shorter fatty acyl chains. This finding demonstrates fatty acids as molecular determinants of APL that regulate hemostasis. Thus, the molecular species of externalized APL during platelet activation, apoptosis, and energy depletion were characterized, and their ability to support coagulation revealed. The findings have therapeutic implications for bleeding disorders and transfusion therapy. The assay could be applied to other cell events characterized by APL externalization, including cell division and vesiculation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Daleke DL. Phospholipid flippases. J Biol Chem. 2007;282(2):821–825. - PubMed
-
- Heemskerk JW, Bevers EM, Lindhout T. Platelet activation and blood coagulation. Thromb Haemost. 2002;88(2):186–193. - PubMed
-
- Daleke DL. Regulation of transbilayer plasma membrane phospholipid asymmetry. J Lipid Res. 2003;44(2):233–242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
