

UBIAD1-mediated vitamin K2 synthesis is required for vascular endothelial cell survival and development
- PMID: 23533172
- PMCID: PMC3621489
- DOI: 10.1242/dev.093112
UBIAD1-mediated vitamin K2 synthesis is required for vascular endothelial cell survival and development
Abstract
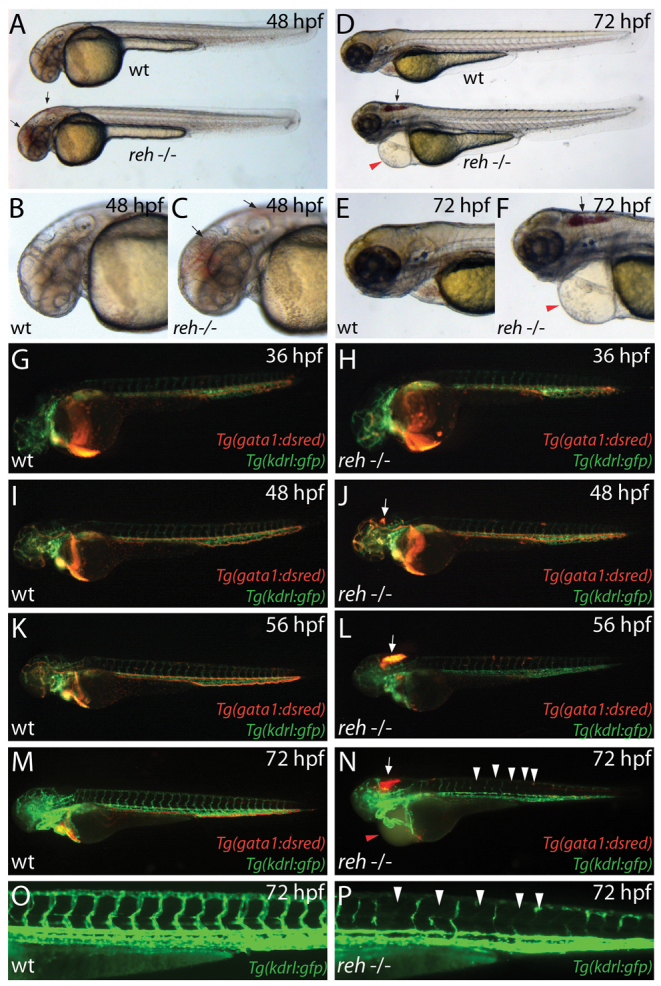
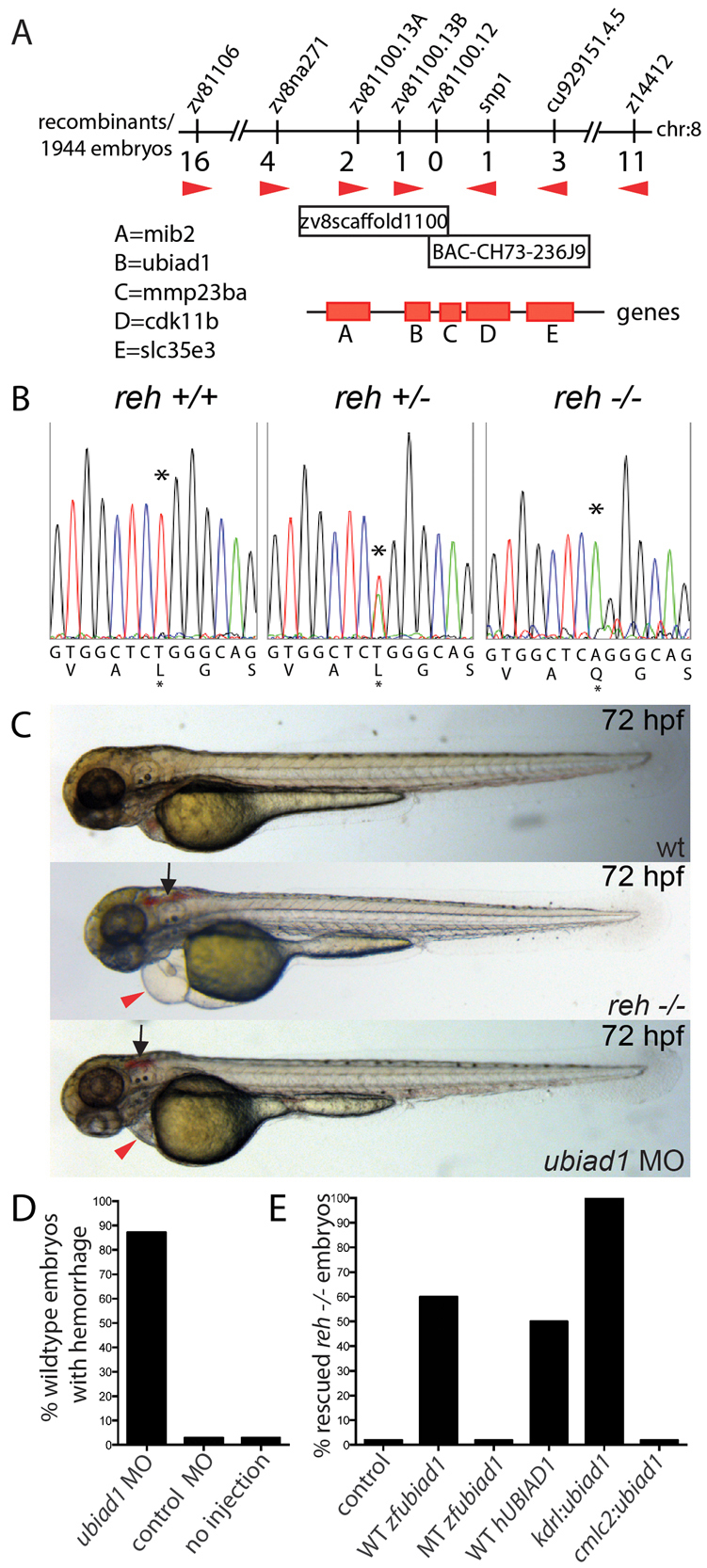
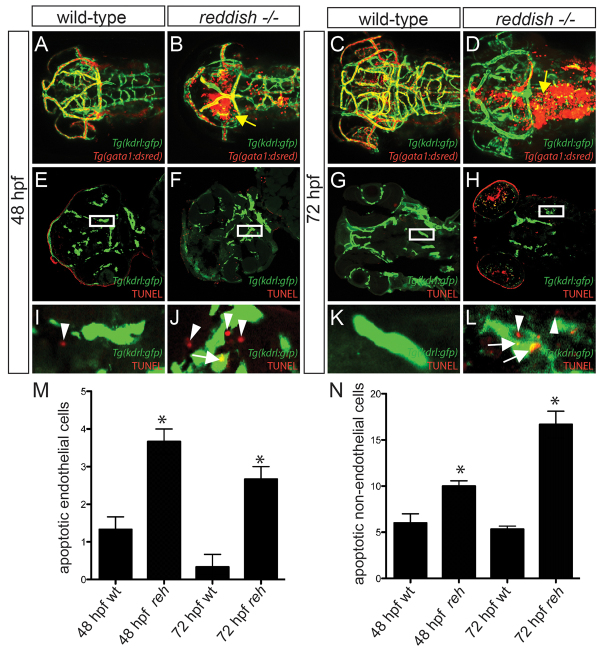
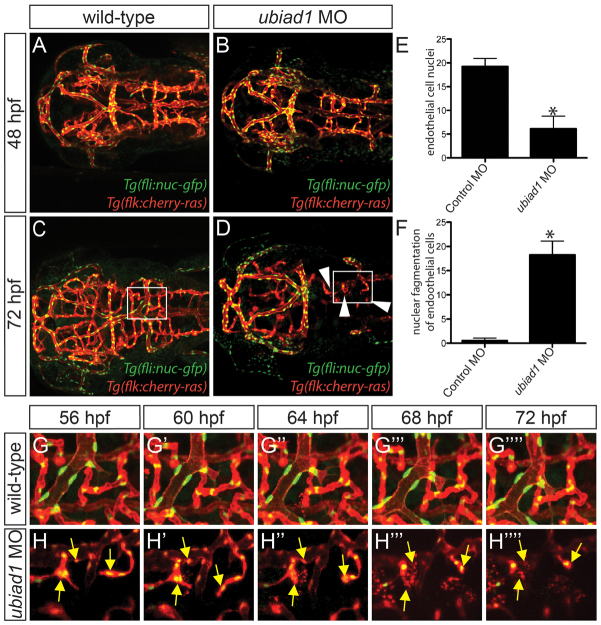
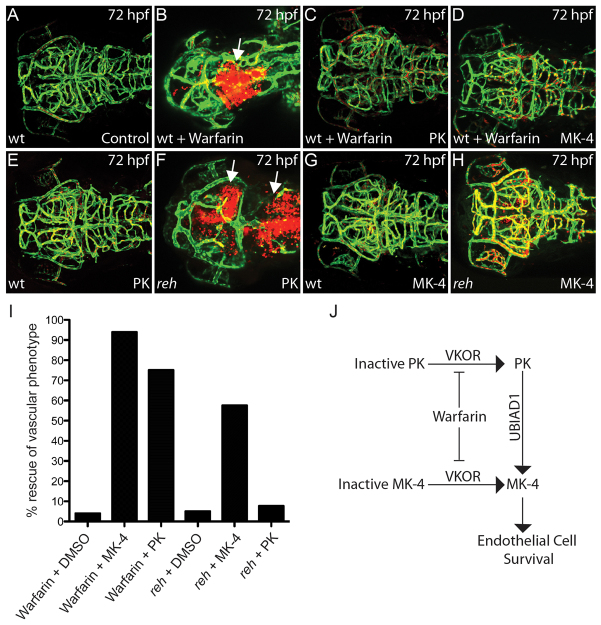
Multi-organ animals, such as vertebrates, require the development of a closed vascular system to ensure the delivery of nutrients to, and the transport of waste from, their organs. As a result, an organized vascular network that is optimal for tissue perfusion is created through not only the generation of new blood vessels but also the remodeling and maintenance of endothelial cells via apoptotic and cell survival pathways. Here, we show that UBIAD1, a vitamin K2/menaquinone-4 biosynthetic enzyme, functions cell-autonomously to regulate endothelial cell survival and maintain vascular homeostasis. From a recent vascular transgene-assisted zebrafish forward genetic screen, we have identified a ubiad1 mutant, reddish/reh, which exhibits cardiac edema as well as cranial hemorrhages and vascular degeneration owing to defects in endothelial cell survival. These findings are further bolstered by the expression of UBIAD1 in human umbilical vein endothelial cells and human heart tissue, as well as the rescue of the reh cardiac and vascular phenotypes with either zebrafish or human UBIAD1. Furthermore, we have discovered that vitamin K2, which is synthesized by UBIAD1, can also rescue the reh vascular phenotype but not the reh cardiac phenotype. Additionally, warfarin-treated zebrafish, which have decreased active vitamin K, display similar vascular degeneration as reh mutants, but exhibit normal cardiac function. Overall, these findings reveal an essential role for UBIAD1-generated vitamin K2 to maintain endothelial cell survival and overall vascular homeostasis; however, an alternative UBIAD1/vitamin K-independent pathway may regulate cardiac function.
Figures
Similar articles
-
Vitamin K2 biosynthetic enzyme, UBIAD1 is essential for embryonic development of mice.PLoS One. 2014 Aug 15;9(8):e104078. doi: 10.1371/journal.pone.0104078. eCollection 2014. PLoS One. 2014. PMID: 25127365 Free PMC article.
-
Functional characterization of the vitamin K2 biosynthetic enzyme UBIAD1.PLoS One. 2015 Apr 15;10(4):e0125737. doi: 10.1371/journal.pone.0125737. eCollection 2015. PLoS One. 2015. PMID: 25874989 Free PMC article.
-
Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme.Nature. 2010 Nov 4;468(7320):117-21. doi: 10.1038/nature09464. Epub 2010 Oct 17. Nature. 2010. PMID: 20953171
-
Recent trends in the metabolism and cell biology of vitamin K with special reference to vitamin K cycling and MK-4 biosynthesis.J Lipid Res. 2014 Mar;55(3):345-62. doi: 10.1194/jlr.R045559. Epub 2014 Jan 31. J Lipid Res. 2014. PMID: 24489112 Free PMC article. Review.
-
[Biological significance and metabolic activation of vitamin K].Yakugaku Zasshi. 2013;133(12):1337-41. doi: 10.1248/yakushi.13-00228-1. Yakugaku Zasshi. 2013. PMID: 24292181 Review. Japanese.
Cited by
-
Animal Models of Coenzyme Q Deficiency: Mechanistic and Translational Learnings.Antioxidants (Basel). 2021 Oct 26;10(11):1687. doi: 10.3390/antiox10111687. Antioxidants (Basel). 2021. PMID: 34829558 Free PMC article. Review.
-
Protective effect of VK2 on glucocorticoid-treated MC3T3-E1 cells.Int J Mol Med. 2017 Jan;39(1):160-166. doi: 10.3892/ijmm.2016.2817. Epub 2016 Dec 1. Int J Mol Med. 2017. PMID: 27909721 Free PMC article.
-
Tissue Concentrations of Vitamin K and Expression of Key Enzymes of Vitamin K Metabolism Are Influenced by Sex and Diet but Not Housing in C57Bl6 Mice.J Nutr. 2016 Aug;146(8):1521-7. doi: 10.3945/jn.116.233130. Epub 2016 Jul 6. J Nutr. 2016. PMID: 27385762 Free PMC article.
-
The atypical Rho GTPase, RhoU, regulates cell-adhesion molecules during cardiac morphogenesis.Dev Biol. 2014 May 15;389(2):182-91. doi: 10.1016/j.ydbio.2014.02.014. Epub 2014 Mar 7. Dev Biol. 2014. PMID: 24607366 Free PMC article.
-
Methods for Structural and Functional Analyses of Intramembrane Prenyltransferases in the UbiA Superfamily.Methods Enzymol. 2017;584:309-347. doi: 10.1016/bs.mie.2016.10.032. Epub 2016 Dec 7. Methods Enzymol. 2017. PMID: 28065269 Free PMC article.
References
-
- Berridge M. J., Lipp P., Bootman M. D. (2000). The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 1, 11–21 - PubMed
-
- Boehm R., Sommer S., Severin K., Li S. M., Heide L. (2000). Active expression of the ubiA gene from E. coli in tobacco: influence of plant ER-specific signal peptides on the expression of a membrane-bound prenyltransferase in plant cells. Transgenic Res. 9, 477–486 - PubMed
-
- Bräuer L., Brandt W., Wessjohann L. A. (2004). Modeling the E. coli 4-hydroxybenzoic acid oligoprenyltransferase (ubiA transferase) and characterization of potential active sites. J. Mol. Model. 10, 317–327 - PubMed
-
- Bräuer L., Brandt W., Schulze D., Zakharova S., Wessjohann L. (2008). A structural model of the membrane-bound aromatic prenyltransferase UbiA from E. coli. ChemBioChem 9, 982–992 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
