

The human CD8β M-4 isoform dominant in effector memory T cells has distinct cytoplasmic motifs that confer unique properties
- PMID: 23533620
- PMCID: PMC3606432
- DOI: 10.1371/journal.pone.0059374
The human CD8β M-4 isoform dominant in effector memory T cells has distinct cytoplasmic motifs that confer unique properties
Abstract
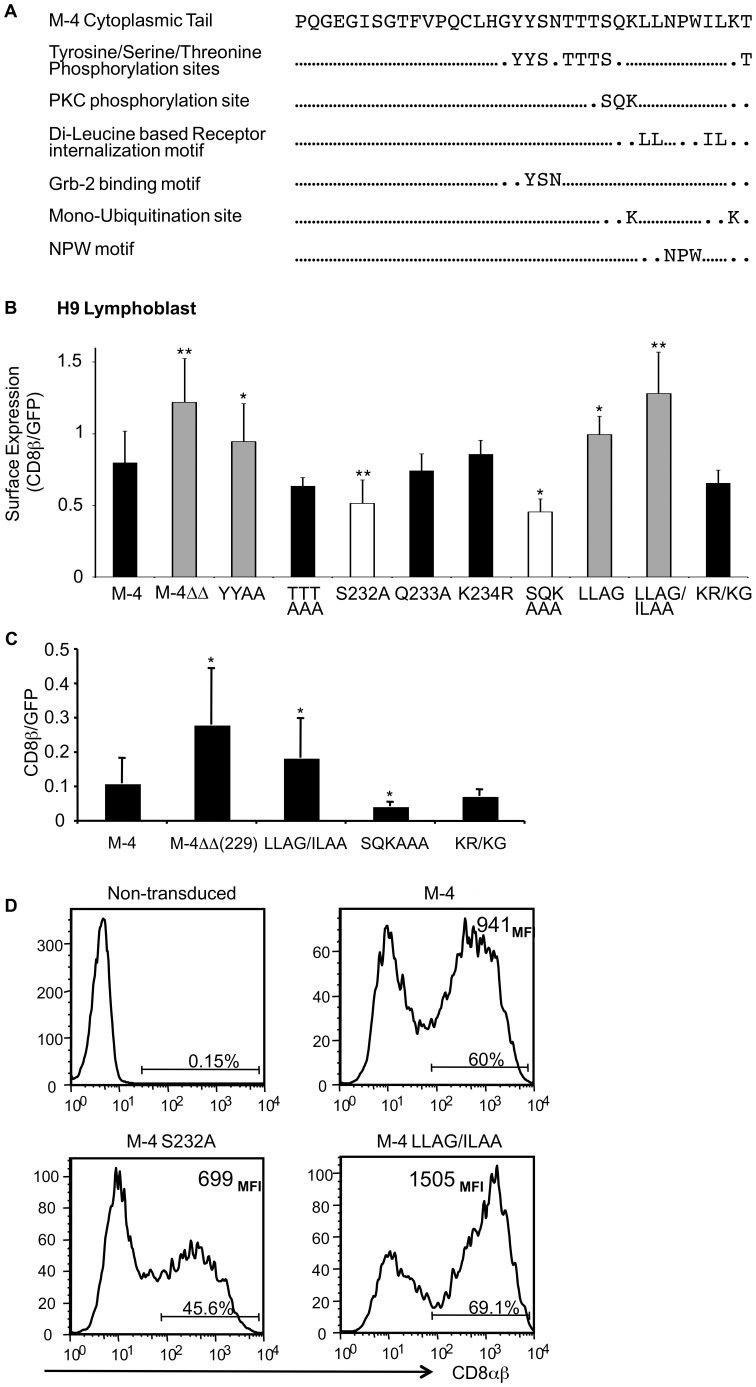
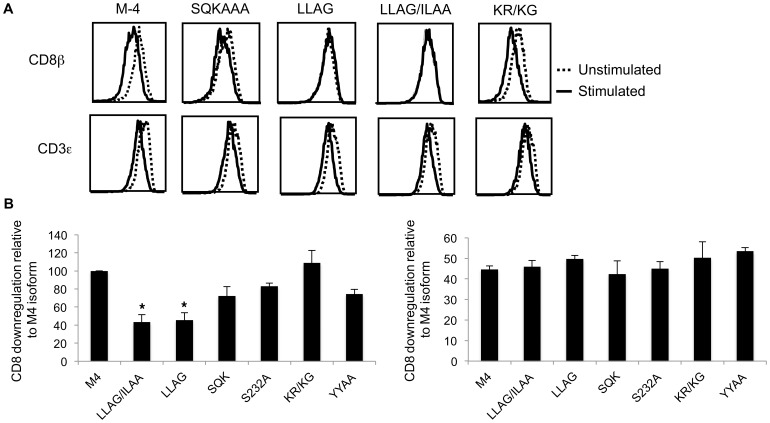
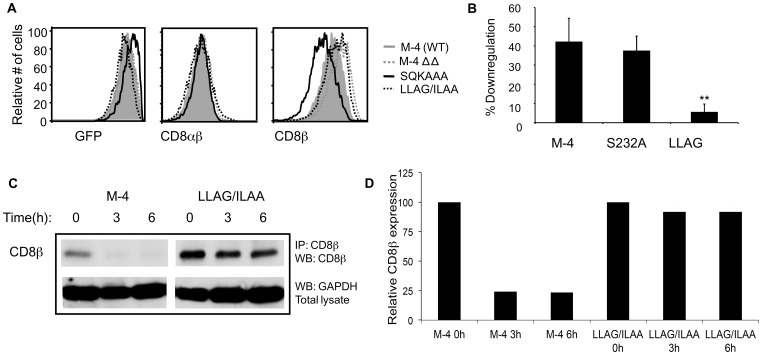
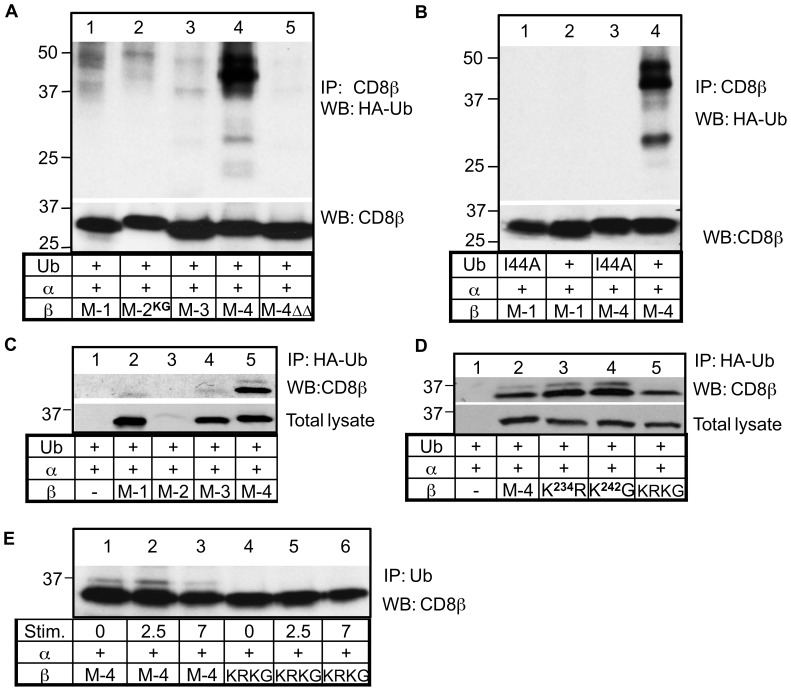
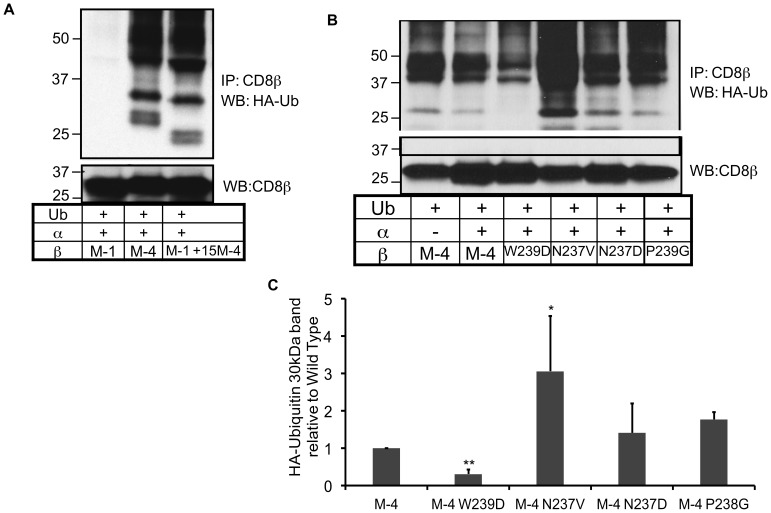
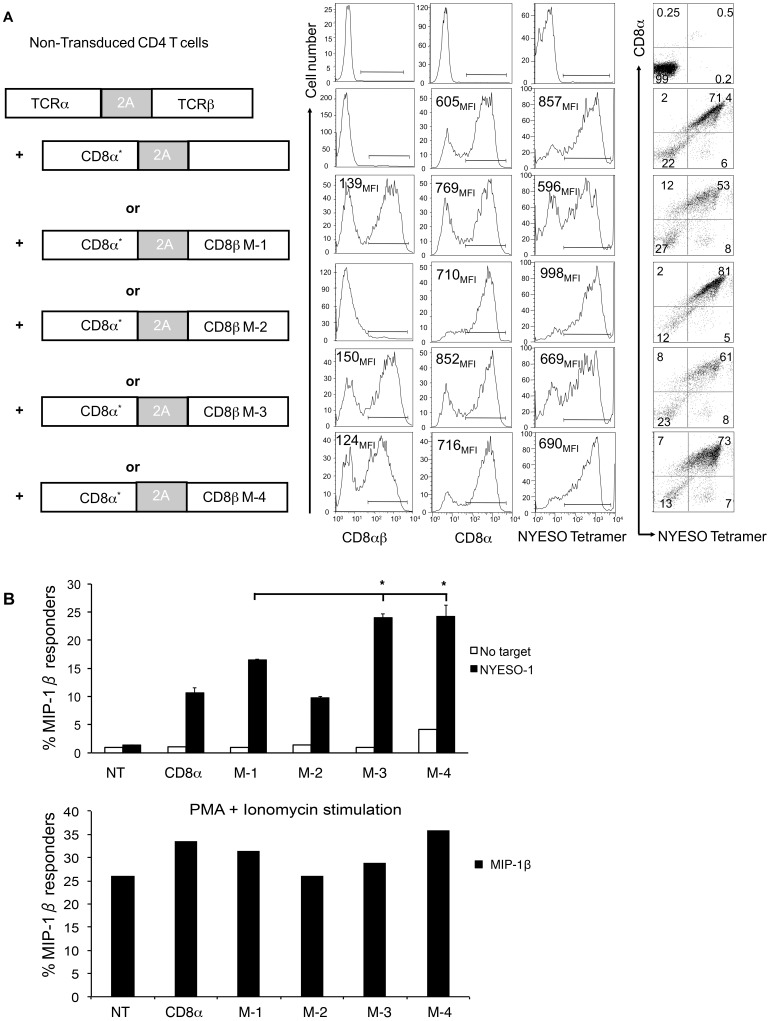
The CD8 co-receptor influences T cell recognition and responses in both anti-tumor and anti-viral immunity. During evolution in the ancestor of humans and chimpanzees, the CD8B gene acquired two additional exons. As a result, in humans, there are four CD8β splice variants (M1 to M4) that differ in their cytoplasmic tails. The M-1 isoform which is the equivalent of murine CD8β, is predominantly expressed in naïve T cells, whereas, the M-4 isoform is predominantly expressed in effector memory T cells. The characteristics of the M-4 isoform conferred by its unique 36 amino acid cytoplasmic tail are not known. In this study, we identified a dihydrophobic leucine-based receptor internalization motif in the cytoplasmic tail of M-4 that regulated its cell surface expression and downregulation after activation. Further the M-4 cytoplasmic tail was able to associate with ubiquitinated targets in 293T cells and mutations in the amino acids NPW, a potential EH domain binding site, either enhanced or inhibited the interaction. In addition, the M-4 tail was itself mono-ubiquitinated on a lysine residue in both 293T cells and a human T cell line. When peripheral blood human T cells expressed CD8αβ M-4, the frequency of MIP-1β secreting cells responding to antigen presenting cells was two-fold higher as compared to CD8αβ M-1 expressing T cells. Thus, the cytoplasmic tail of the CD8β M-4 isoform has unique characteristics, which likely contributed to its selective expression and function in human effector memory T cells.
Conflict of interest statement
Figures
Similar articles
-
In vitro IL-15-activated human naïve CD8+ T cells down-modulate the CD8β chain and become CD8αα T cells.Front Immunol. 2024 Jun 5;15:1252439. doi: 10.3389/fimmu.2024.1252439. eCollection 2024. Front Immunol. 2024. PMID: 38903513 Free PMC article.
-
Differential expression of the human CD8beta splice variants and regulation of the M-2 isoform by ubiquitination.J Immunol. 2008 Jun 1;180(11):7431-42. doi: 10.4049/jimmunol.180.11.7431. J Immunol. 2008. PMID: 18490743
-
Mapping the binding site on CD8 beta for MHC class I reveals mutants with enhanced binding.J Immunol. 2006 Sep 15;177(6):3930-8. doi: 10.4049/jimmunol.177.6.3930. J Immunol. 2006. PMID: 16951356
-
Chicken CD4, CD8alphabeta, and CD8alphaalpha T cell co-receptor molecules.Poult Sci. 1998 Dec;77(12):1858-73. doi: 10.1093/ps/77.12.1858. Poult Sci. 1998. PMID: 9872590 Review.
-
Molecular interactions of coreceptor CD8 and MHC class I: the molecular basis for functional coordination with the T-cell receptor.Immunol Today. 2000 Dec;21(12):630-6. doi: 10.1016/s0167-5699(00)01750-3. Immunol Today. 2000. PMID: 11114424 Review.
Cited by
-
In vitro IL-15-activated human naïve CD8+ T cells down-modulate the CD8β chain and become CD8αα T cells.Front Immunol. 2024 Jun 5;15:1252439. doi: 10.3389/fimmu.2024.1252439. eCollection 2024. Front Immunol. 2024. PMID: 38903513 Free PMC article.
-
Development of T-cell immunotherapy for hematopoietic stem cell transplantation recipients at risk of leukemia relapse.Blood. 2018 Jan 4;131(1):108-120. doi: 10.1182/blood-2017-07-791608. Epub 2017 Oct 19. Blood. 2018. PMID: 29051183 Free PMC article.
References
-
- Russell JH, Ley TJ (2002) Lymphocyte-mediated cytotoxicity. Annu Rev Immunol 20: 323–370. - PubMed
-
- Yachi PP, Ampudia J, Zal T, Gascoigne NR (2006) Altered peptide ligands induce delayed CD8-T cell receptor interaction–a role for CD8 in distinguishing antigen quality. Immunity 25: 203–211. - PubMed
-
- Holler PD, Kranz DM (2003) Quantitative analysis of the contribution of TCR/pepMHC affinity and CD8 to T cell activation. Immunity 18: 255–264. - PubMed
-
- Purbhoo MA, Boulter JM, Price DA, Vuidepot AL, Hourigan CS, et al. (2001) The human CD8 coreceptor effects cytotoxic T cell activation and antigen sensitivity primarily by mediating complete phosphorylation of the T cell receptor zeta chain. J Biol Chem 276: 32786–32792. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
