

Tumor suppressor protein Pdcd4 interacts with Daxx and modulates the stability of Daxx and the Hipk2-dependent phosphorylation of p53 at serine 46
- PMID: 23536002
- PMCID: PMC3564021
- DOI: 10.1038/oncsis.2012.37
Tumor suppressor protein Pdcd4 interacts with Daxx and modulates the stability of Daxx and the Hipk2-dependent phosphorylation of p53 at serine 46
Abstract
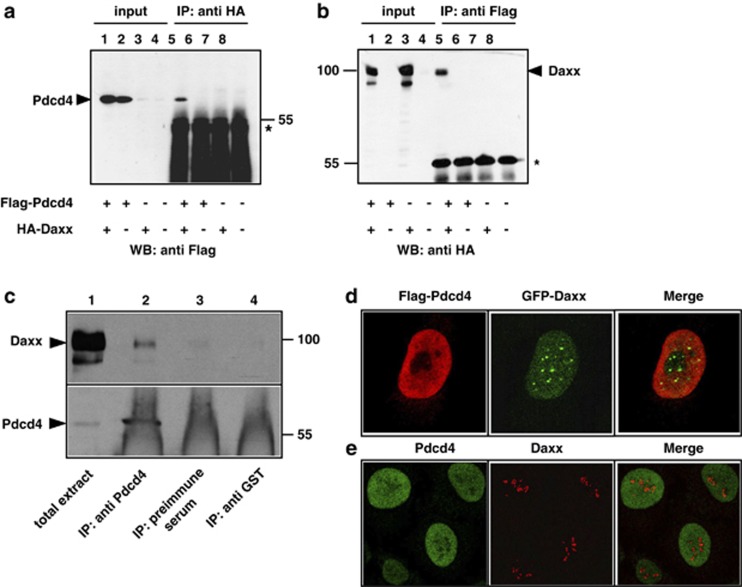
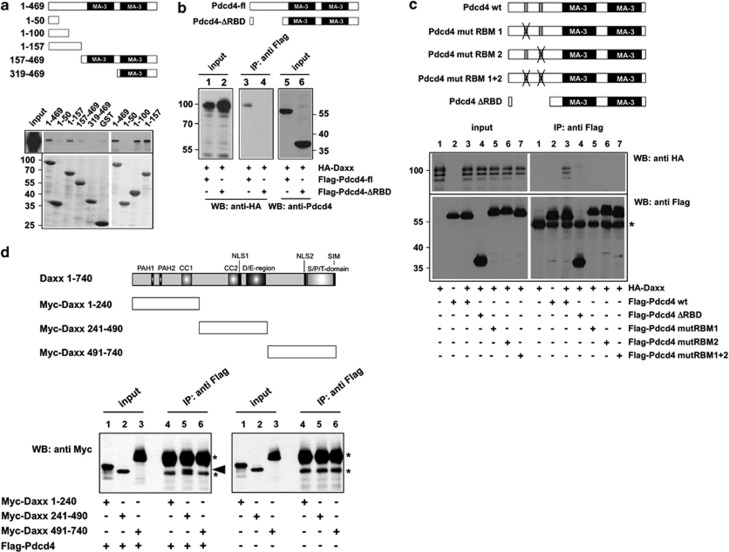
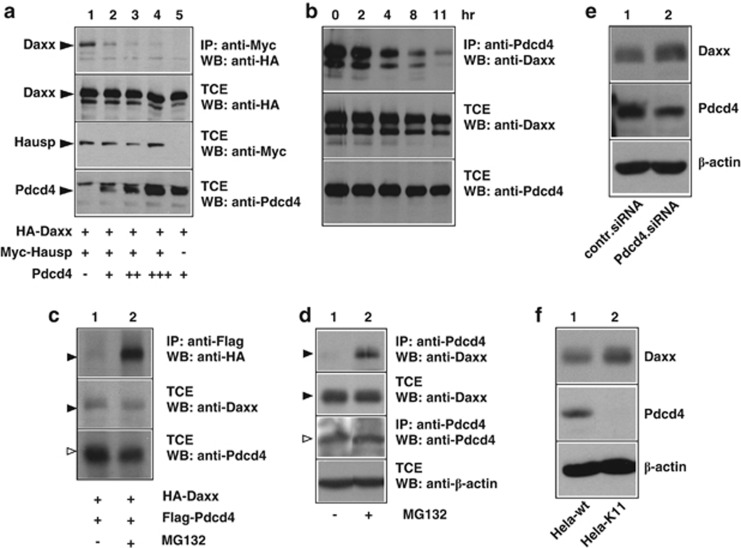
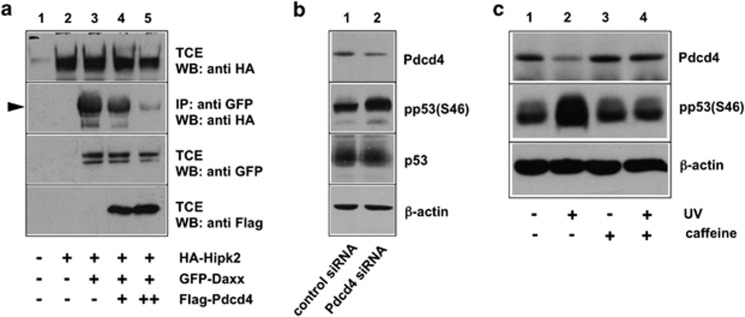
The tumor suppressor protein Pdcd4 is a nuclear/cytoplasmic shuttling protein that has been implicated in the development of several types of human cancer. In the nucleus, Pdcd4 affects the transcription of specific genes by modulating the activity of several transcription factors. We have identified the Daxx protein as a novel interaction partner of Pdcd4. Daxx is a scaffold protein with roles in diverse processes, including transcriptional regulation, DNA-damage signaling, apoptosis and chromatin remodeling. We show that the interaction of both proteins is mediated by the N-terminal domain of Pdcd4 and the central part of Daxx, and that binding to Pdcd4 stimulates the degradation of Daxx, presumably by disrupting the interaction of Daxx with the de-ubiquitinylating enzyme Hausp. Daxx has previously been shown to serve as a scaffold for protein kinase Hipk2 and tumor suppressor protein p53 and to stimulate the phosphorylation of p53 at serine 46 (Ser-46) in response to genotoxic stress. We show that Pdcd4 also disrupts the Daxx-Hipk2 interaction and inhibits the phosphorylation of p53. We also show that ultraviolet irradiation decreases the expression of Pdcd4. Taken together, our results support a model in which Pdcd4 serves to suppress the phosphorylation of p53 in the absence of DNA damage, while the suppressive effect of Pdcd4 is abrogated after DNA damage owing to the decrease of Pdcd4. Overall, our data demonstrate that Pdcd4 is a novel modulator of Daxx function and provide evidence for a role of Pdcd4 in restraining p53 activity in unstressed cells.
Figures
References
-
- Shibahara K, Asano M, Ishida Y, Aoki T, Koike T, Honjo T. Isolation of a novel mouse gene MA-3 that is induced upon programmed cell death. Gene. 1995;166:297–301. - PubMed
-
- Jansen AP, Camalier CE, Colburn NH. Epidermal expression of the translation inhibitor programmed cell death 4 suppresses tumorigenesis. Cancer Res. 2005;65:6034–6041. - PubMed
-
- Chen Y, Knosel T, Kristiansen G, Pietas A, Garber ME, Matsuhashi S, et al. Loss of PDCD4 expression in human lung cancer correlates with tumour progression and prognosis. J Pathol. 2003;200:640–646. - PubMed
-
- Afonja O, Juste D, Das S, Matsuhashi S, Samuels HH. Induction of PDCD4 tumor suppressor gene expression by RAR agonists, antiestrogen and HER-2/neu antagonist in breast cancer cells. Evidence for a role in apoptosis. Oncogene. 2004;23:8135–8145. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
