

ACR-12 ionotropic acetylcholine receptor complexes regulate inhibitory motor neuron activity in Caenorhabditis elegans
- PMID: 23536067
- PMCID: PMC3645261
- DOI: 10.1523/JNEUROSCI.4384-12.2013
ACR-12 ionotropic acetylcholine receptor complexes regulate inhibitory motor neuron activity in Caenorhabditis elegans
Abstract
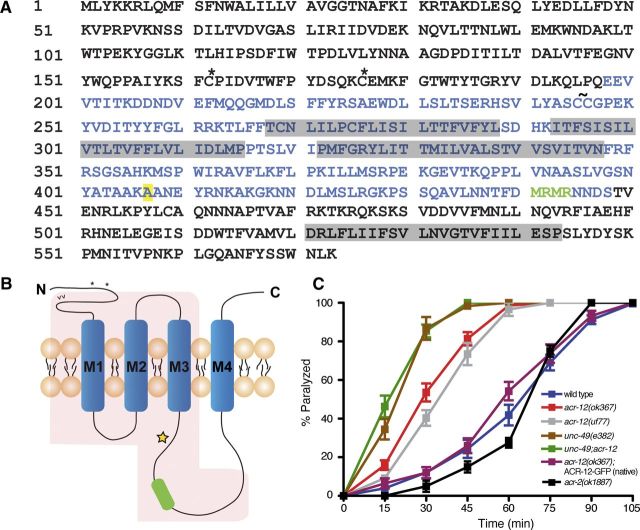
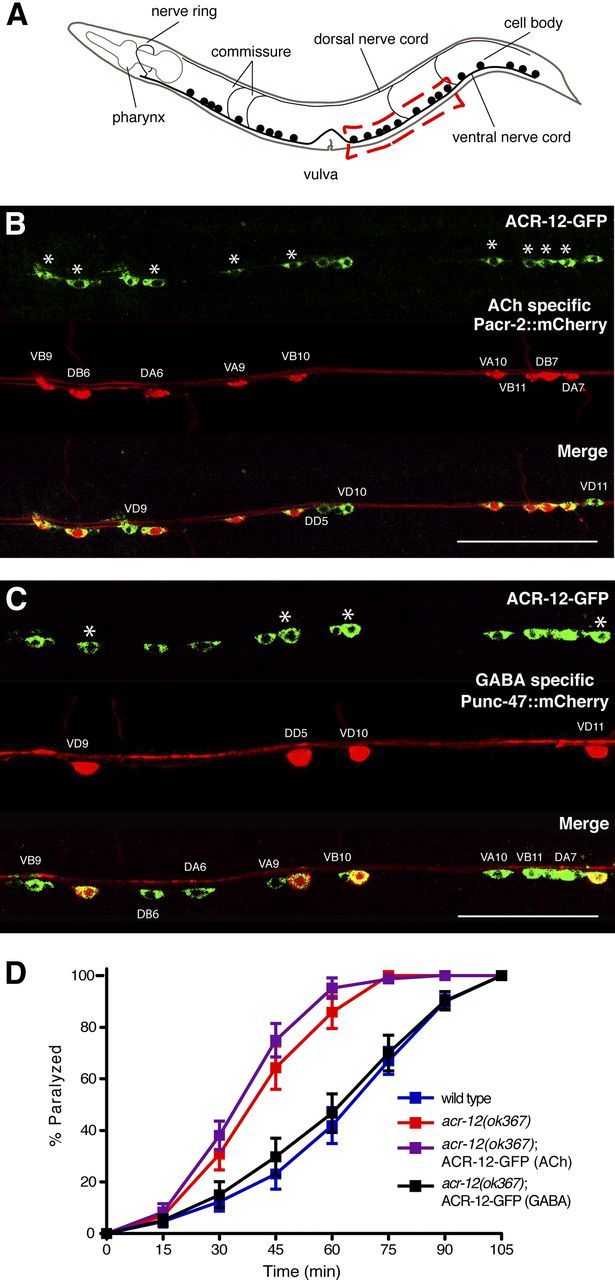
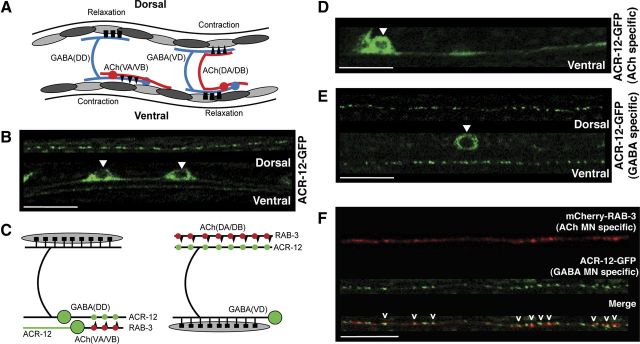
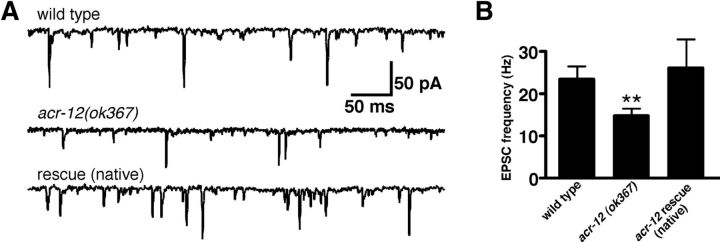
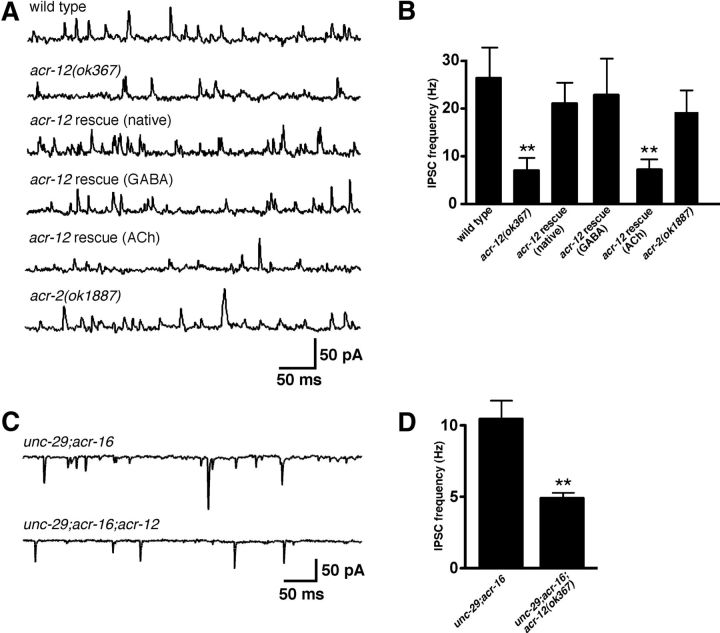
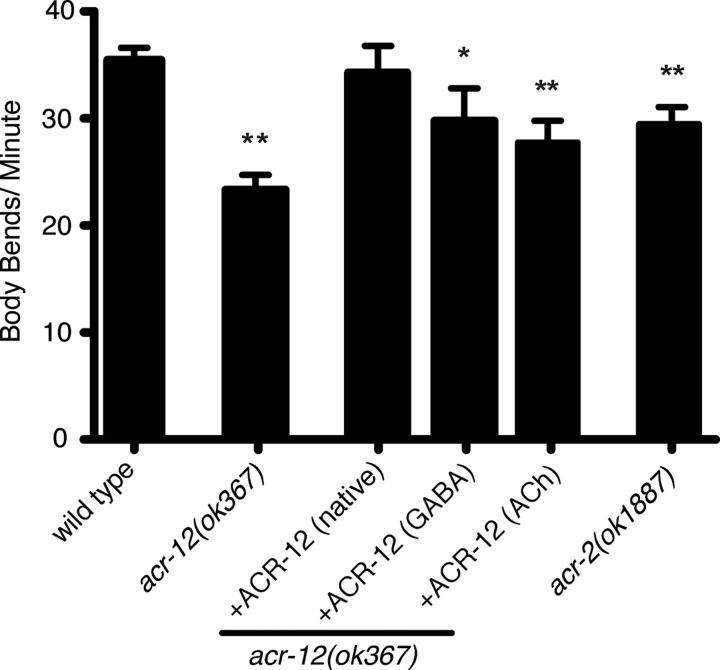
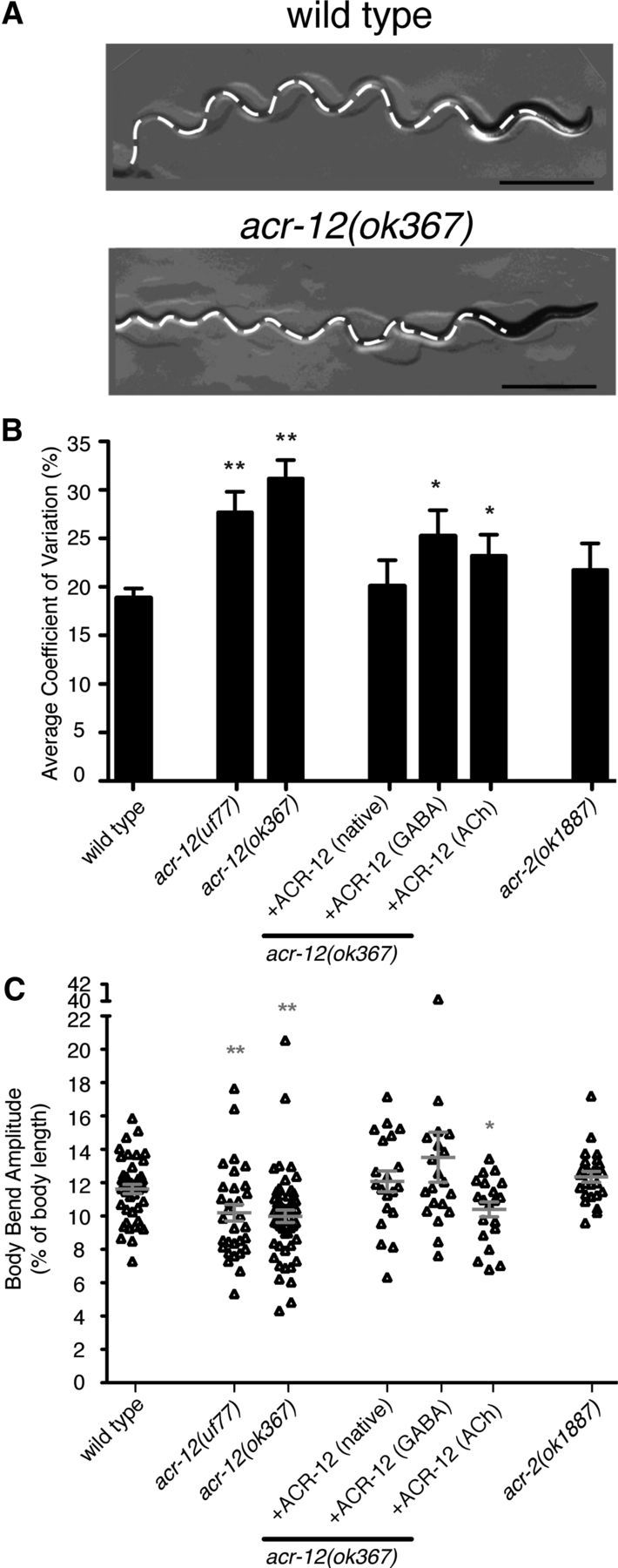
Heterogeneity in the composition of neurotransmitter receptors is thought to provide functional diversity that may be important in patterning neural activity and shaping behavior (Dani and Bertrand, 2007; Sassoè-Pognetto, 2011). However, this idea has remained difficult to evaluate directly because of the complexity of neuronal connectivity patterns and uncertainty about the molecular composition of specific receptor types in vivo. Here we dissect how molecular diversity across receptor types contributes to the coordinated activity of excitatory and inhibitory motor neurons in the nematode Caenorhabditis elegans. We show that excitatory and inhibitory motor neurons express distinct populations of ionotropic acetylcholine receptors (iAChRs) requiring the ACR-12 subunit. The activity level of excitatory motor neurons is influenced through activation of nonsynaptic iAChRs (Jospin et al., 2009; Barbagallo et al., 2010). In contrast, synaptic coupling of excitatory and inhibitory motor neurons is achieved through a second population of iAChRs specifically localized at postsynaptic sites on inhibitory motor neurons. Loss of ACR-12 iAChRs from inhibitory motor neurons leads to reduced synaptic drive, decreased inhibitory neuromuscular signaling, and variability in the sinusoidal motor pattern. Our results provide new insights into mechanisms that establish appropriately balanced excitation and inhibition in the generation of a rhythmic motor behavior and reveal functionally diverse roles for iAChR-mediated signaling in this process.
Figures
References
-
- Cinar H, Keles S, Jin Y. Expression profiling of GABAergic motor neurons in Caenorhabditis elegans. Curr Biol. 2005;15:340–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources