

Conditional disruption of calpain in the CNS alters dendrite morphology, impairs LTP, and promotes neuronal survival following injury
- PMID: 23536090
- PMCID: PMC4153560
- DOI: 10.1523/JNEUROSCI.4247-12.2013
Conditional disruption of calpain in the CNS alters dendrite morphology, impairs LTP, and promotes neuronal survival following injury
Abstract
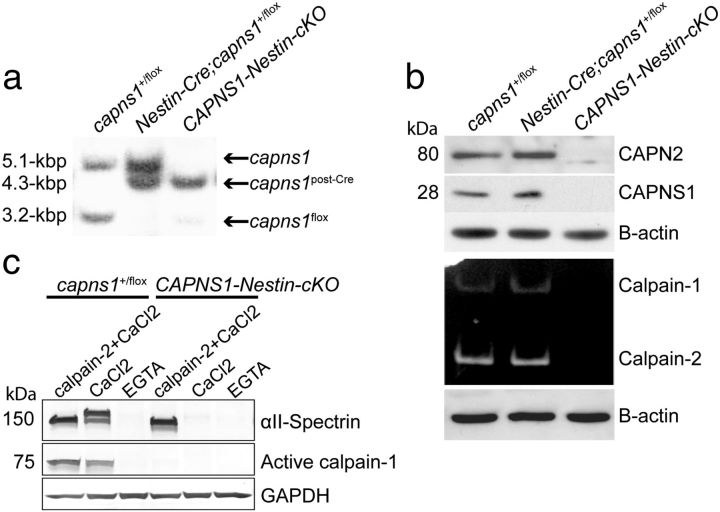
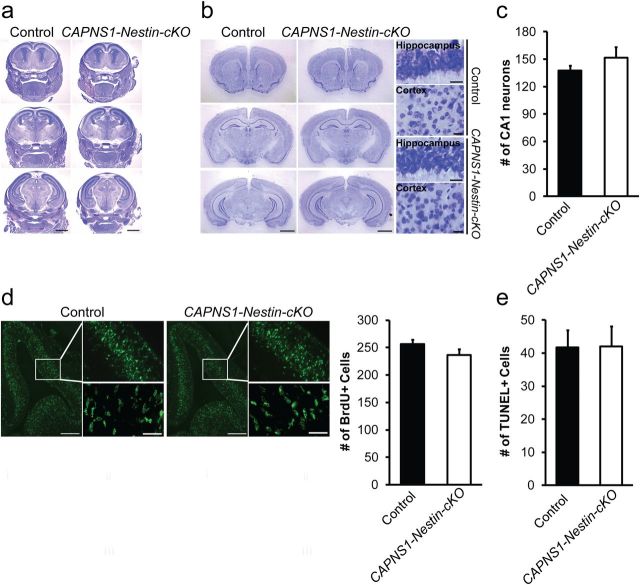
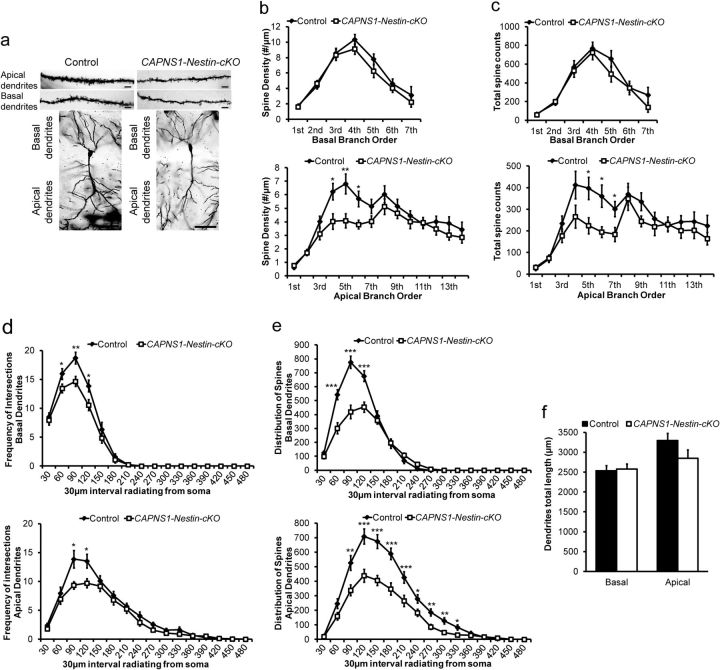
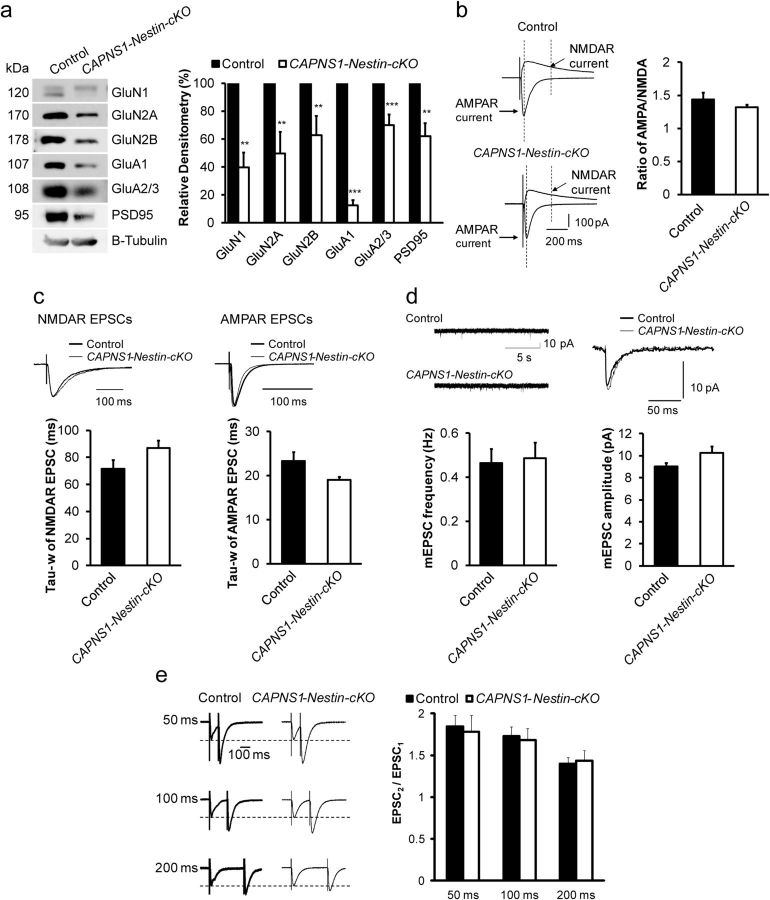
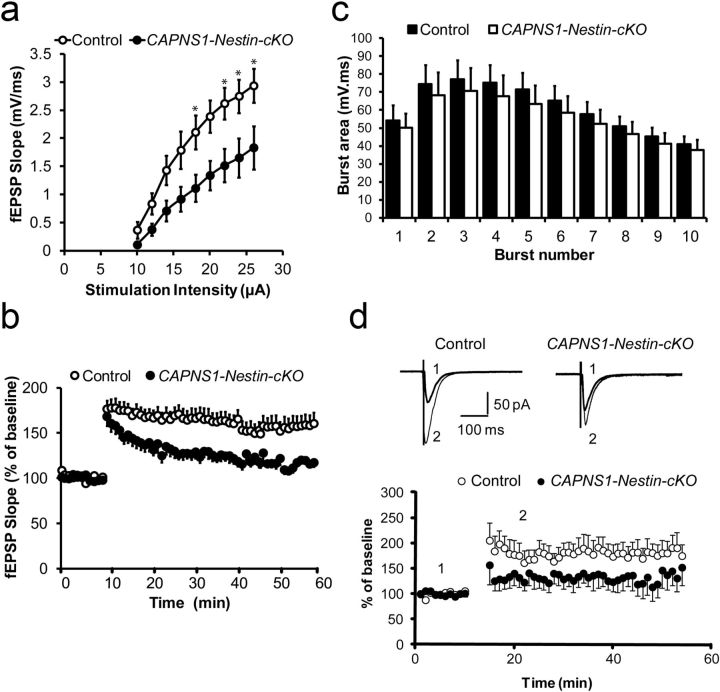
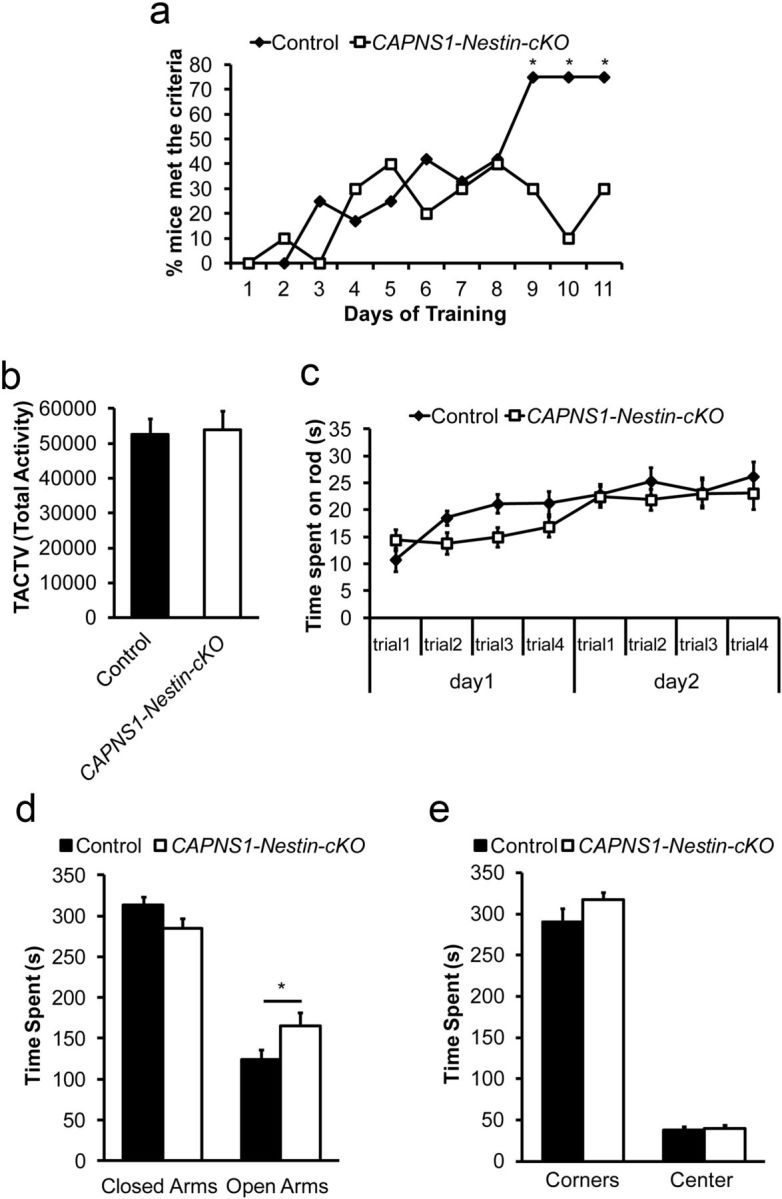
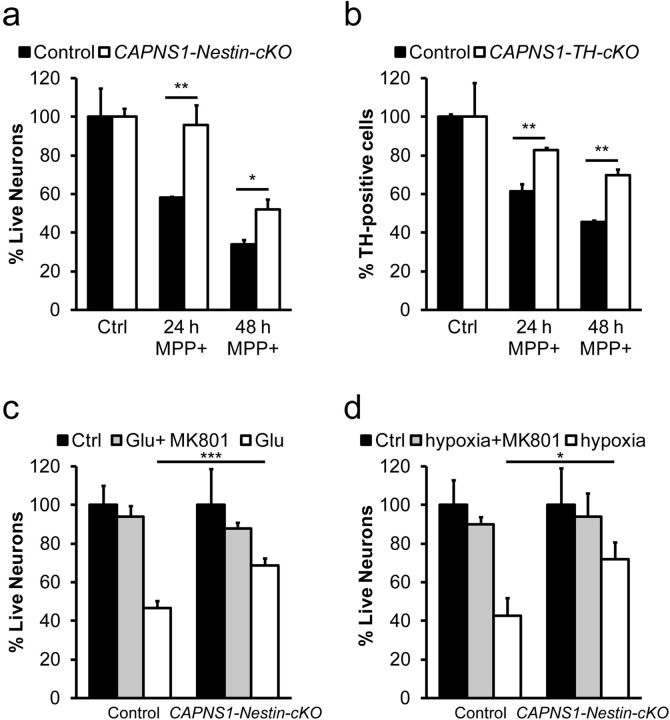
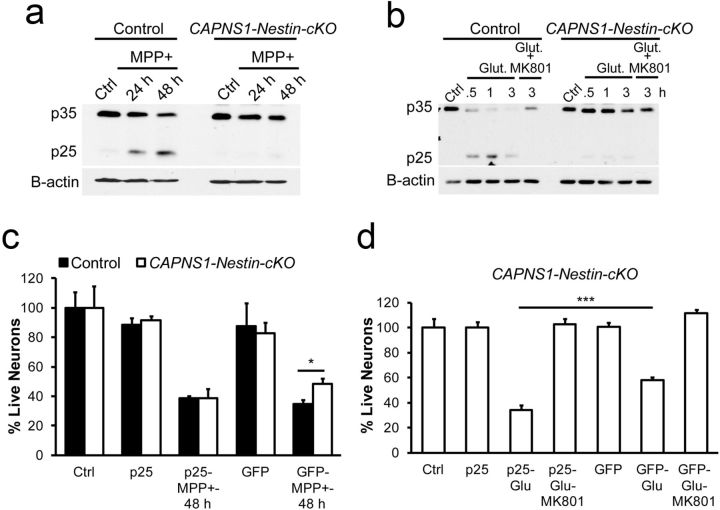
Ubiquitous classical (typical) calpains, calpain-1 and calpain-2, are Ca(+2)-dependent cysteine proteases, which have been associated with numerous physiological and pathological cellular functions. However, a clear understanding of the role of calpains in the CNS has been hampered by the lack of appropriate deletion paradigms in the brain. In this study, we describe a unique model of conditional deletion of both calpain-1 and calpain-2 activities in mouse brain, which more definitively assesses the role of these ubiquitous proteases in brain development/function and pathology. Surprisingly, we show that these calpains are not critical for gross CNS development. However, calpain-1/calpain-2 loss leads to reduced dendritic branching complexity and spine density deficits associated with major deterioration in hippocampal long-term potentiation and spatial memory. Moreover, calpain-1/calpain-2-deficient neurons were significantly resistant to injury induced by excitotoxic stress or mitochondrial toxicity. Examination of downstream target showed that the conversion of the Cdk5 activator, p35, to pathogenic p25 form, occurred only in the presence of calpain and that it played a major role in calpain-mediated neuronal death. These findings unequivocally establish two central roles of calpain-1/calpain-2 in CNS function in plasticity and neuronal death.
Figures
References
-
- Bai G, Hoffman PW. Transcriptional regulation of NMDA receptor expression. In: Van Dongen AM, editor. Boca Raton, FL: CRC; 2009. - PubMed
-
- Bartus RT, Dean RL, Cavanaugh K, Eveleth D, Carriero DL, Lynch G. Time-related neuronal changes following middle cerebral artery occlusion: implications for therapeutic intervention and the role of calpain. J Cereb Blood Flow Metab. 1995;15:969–979. - PubMed
-
- Baudry M, Lynch G. Remembrance of arguments past: how well is the glutamate receptor hypothesis of LTP holding up after 20 years? Neurobiol Learn Mem. 2001;76:284–297. - PubMed
-
- Bednarski E, Vanderklish P, Gall C, Saido TC, Bahr BA, Lynch G. Translational suppression of calpain I reduces NMDA-induced spectrin proteolysis and pathophysiology in cultured hippocampal slices. Brain Res. 1995;694:147–157. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials