

Tracking the development of the petaloid fertile stamen in Canna indica: insights into the origin of androecial petaloidy in the Zingiberales
- PMID: 23539493
- PMCID: PMC3608240
- DOI: 10.1093/aobpla/plt009
Tracking the development of the petaloid fertile stamen in Canna indica: insights into the origin of androecial petaloidy in the Zingiberales
Abstract
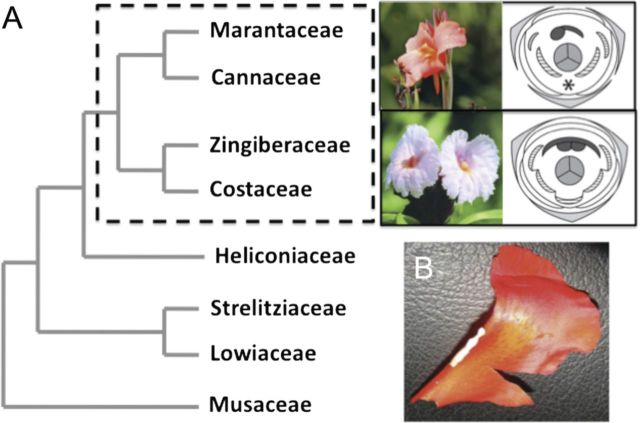
Flowers of the order Zingiberales demonstrate a remarkable trend of reduction in the number of fertile stamens; from five or six fertile, filamentous stamens bearing two thecae each in Musaceae and Strelitziaceae to just a single petaloid stamen bearing a single theca in Cannaceae and Marantaceae. As one progresses from ancestral to derived floral forms, 5-6 fertile stamens are replaced by 4-5 petaloid staminodes. In Cannaceae and Costaceae, all members of the androecial whorls exhibit petaloidy, including the fertile stamen. In Costaceae, a single fertile stamen develops two thecae embedded on a broad petaloid appendage, while in Cannaceae the single fertile stamen is further reduced to a single theca with a prominent, expanded petaloid appendage. Whether petaloidy of the fertile stamen is a synapomorphy of the entire ginger clade (including Cannaceae, Costaceae, Zingiberaceae and Marantaceae), or the result of independent convergent evolution in Cannaceae, Costaceae, and some Zingiberaceae, is unclear. We combine a developmental series of the formation of the petaloid fertile stamen in Canna indica with data on the expression of B- and C-class floral organ identity genes to elucidate the organogenetic identity of the petaloid stamen and staminodes. Our data indicate that the single fertile theca in C. indica and its petaloid appendage are derived from one-half of the primordium of a single stamen, with no contribution from the remaining part of the stamen (i.e. the second theca primordium) which aborts early in development. The petaloid appendage expands later, and develops from the position of the filament/connective of the developing theca. Floral identity gene expression shows that petal identity genes (i.e. B-class genes) are expressed in all floral organs studied while C-class gene AG-1 is expressed in an increasing gradient from sepals to gynoecium, and AG-2 is expressed in all floral organs except the petals. The canonical model for molecular specification of floral organ identity is not sufficient to explain petaloidy in the androecial whorl in Canna sp. Further studies understanding the regulation of gene networks are required.
Keywords: Canna; MADS-box genes; Zingiberales; evo-devo; floral development; petaloid stamens; petaloidy.
Figures



References
-
- Bartlett ME, Kirchoff BK, Specht CD. Epi-illumination microscopy coupled to in situ hybridization in non-model species. Development Genes and Evolution. 2008;218:273–279. - PubMed
-
- Bowman JL, Smyth DR, Meyerowitz EM. Genetic interactions among floral homeotic genes of Arabidopsis. Development. 1991;112:1–20. - PubMed
-
- Bremer B, Bremer K, et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Botanical Journal of the Linnean Society. 2009;161:105–121. The Angiosperm Phylogeny Group.
-
- Charlton WA, MacDonald AD, Posluszny U, Wilkins CP. Additions to the technique of epi-illumination light microscopy for the study of floral and vegetative apices. Canadian Journal of Botany. 1989;67:1739–1743.
-
- Coen E, Meyerowitz E. The war of the whorls—genetic interactions controlling flower development. Nature. 1991;353:31–37. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
