

Heme sensor proteins
- PMID: 23539616
- PMCID: PMC3650359
- DOI: 10.1074/jbc.R112.422642
Heme sensor proteins
Abstract
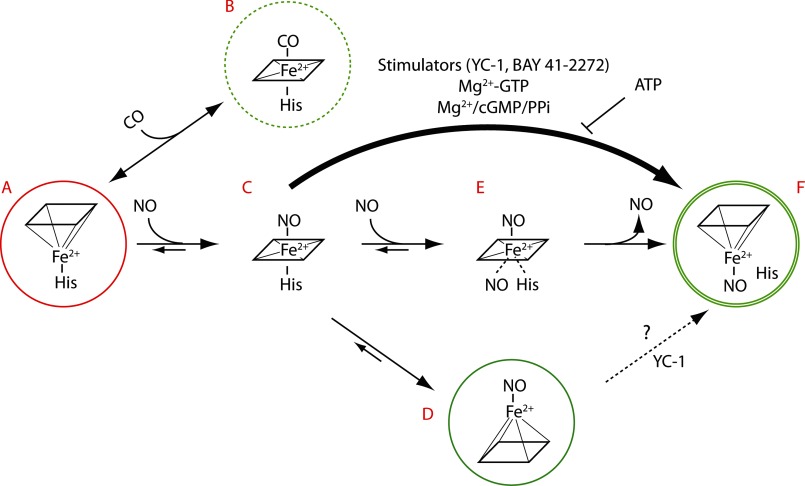
Heme is a prosthetic group best known for roles in oxygen transport, oxidative catalysis, and respiratory electron transport. Recent years have seen the roles of heme extended to sensors of gases such as O2 and NO and cell redox state, and as mediators of cellular responses to changes in intracellular levels of these gases. The importance of heme is further evident from identification of proteins that bind heme reversibly, using it as a signal, e.g. to regulate gene expression in circadian rhythm pathways and control heme synthesis itself. In this minireview, we explore the current knowledge of the diverse roles of heme sensor proteins.
Keywords: Carbon Monoxide; Circadian Rhythms; Cytochromes; Gas Sensors; Gene Regulation; Heme; Nitric Oxide; Nuclear Receptors; Oxygen; Redox Sensor.
Figures


References
-
- Halliwell B., Gutteridge J. M. (1990) Role of free radicals and catalytic metal ions in human disease: an overview. Methods Enzymol. 186, 1–85 - PubMed
-
- Bain D. L., Heneghan A. F., Connaghan-Jones K. D., Miura M. T. (2007) Nuclear receptor structure: implications for function. Annu. Rev. Physiol. 69, 201–220 - PubMed
-
- Kaasik K., Lee C. C. (2004) Reciprocal regulation of haem biosynthesis and the circadian clock in mammals. Nature 430, 467–471 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
