

Elongation factor Ts directly facilitates the formation and disassembly of the Escherichia coli elongation factor Tu·GTP·aminoacyl-tRNA ternary complex
- PMID: 23539628
- PMCID: PMC3650427
- DOI: 10.1074/jbc.M113.460014
Elongation factor Ts directly facilitates the formation and disassembly of the Escherichia coli elongation factor Tu·GTP·aminoacyl-tRNA ternary complex
Abstract
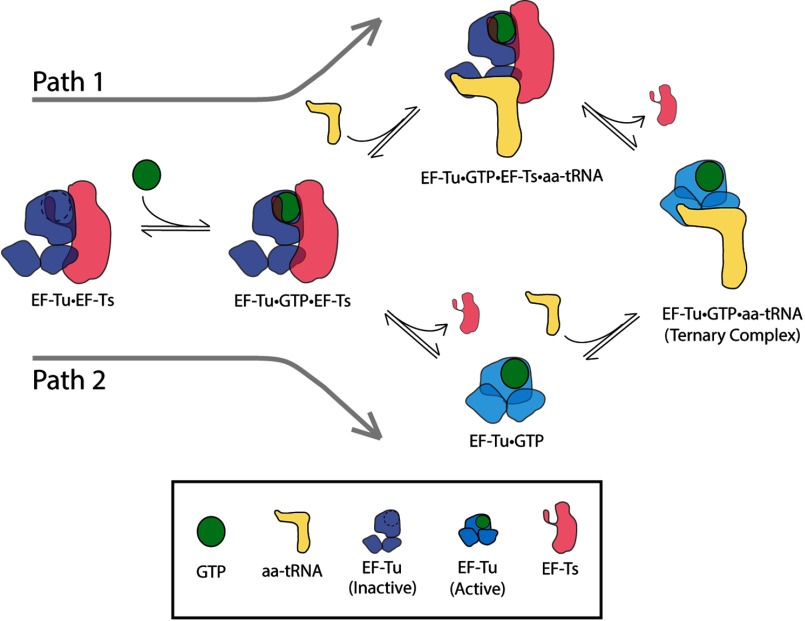
Background: Aminoacyl-tRNA (aa-tRNA) enters the ribosome in a ternary complex with the G-protein elongation factor Tu (EF-Tu) and GTP.
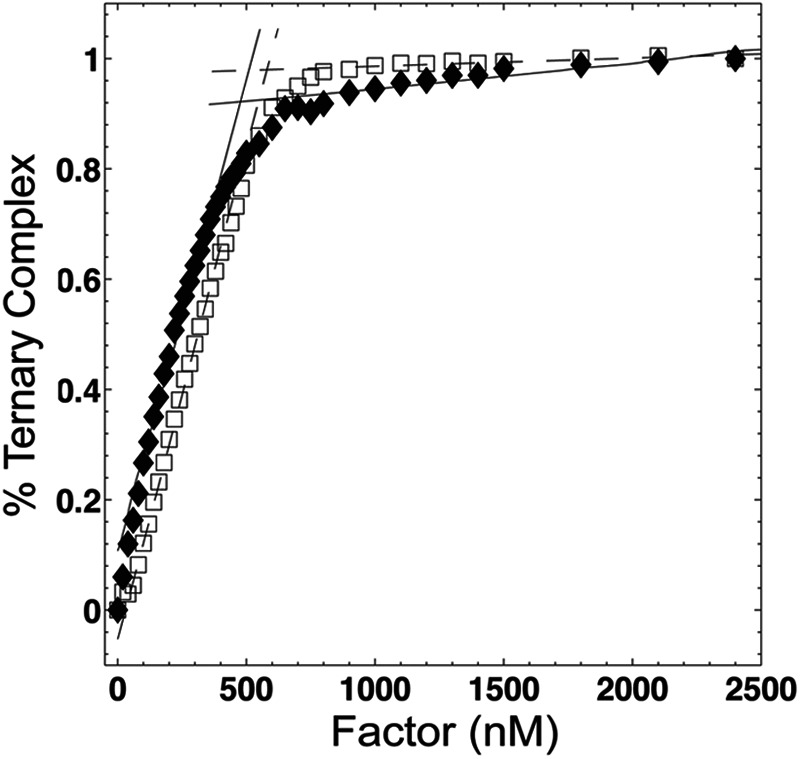
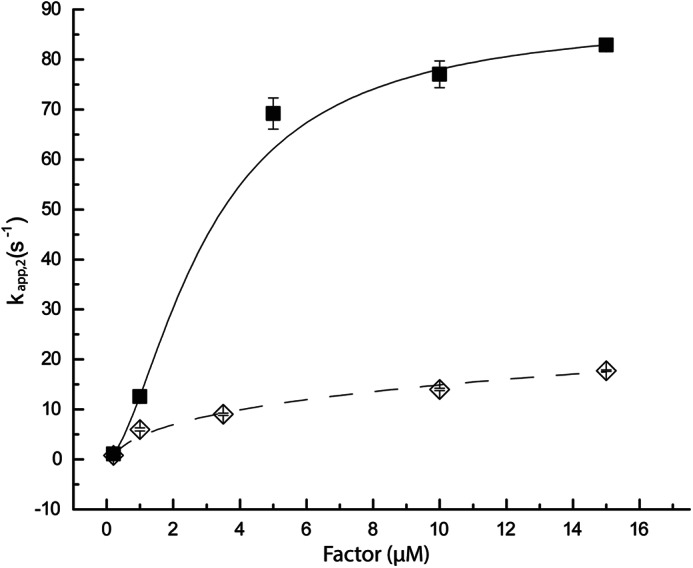
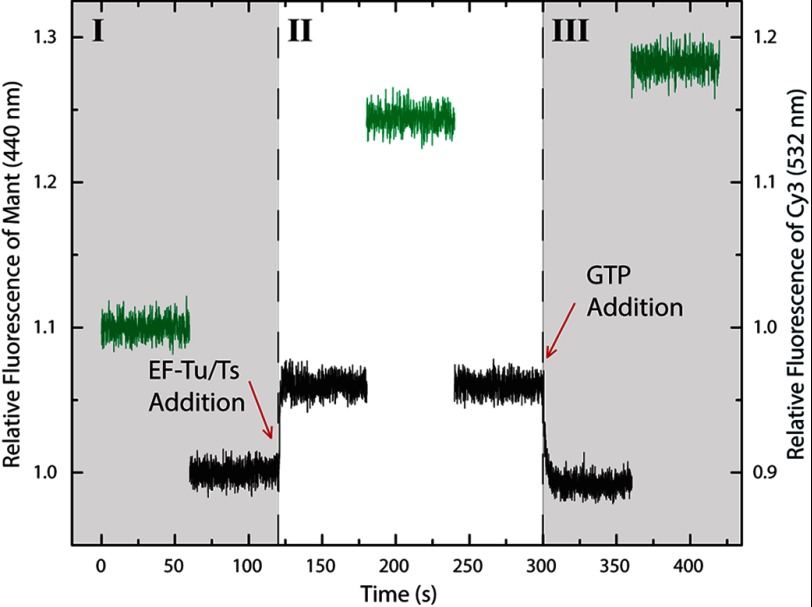
Results: EF-Tu·GTP·aa-tRNA ternary complex formation and decay rates are accelerated in the presence of the nucleotide exchange factor elongation factor Ts (EF-Ts).
Conclusion: EF-Ts directly facilitates the formation and disassociation of ternary complex.
Significance: This system demonstrates a novel function of EF-Ts. Aminoacyl-tRNA enters the translating ribosome in a ternary complex with elongation factor Tu (EF-Tu) and GTP. Here, we describe bulk steady state and pre-steady state fluorescence methods that enabled us to quantitatively explore the kinetic features of Escherichia coli ternary complex formation and decay. The data obtained suggest that both processes are controlled by a nucleotide-dependent, rate-determining conformational change in EF-Tu. Unexpectedly, we found that this conformational change is accelerated by elongation factor Ts (EF-Ts), the guanosine nucleotide exchange factor for EF-Tu. Notably, EF-Ts attenuates the affinity of EF-Tu for GTP and destabilizes ternary complex in the presence of non-hydrolyzable GTP analogs. These results suggest that EF-Ts serves an unanticipated role in the cell of actively regulating the abundance and stability of ternary complex in a manner that contributes to rapid and faithful protein synthesis.
Keywords: Elongation Factor Ts; Elongation Factor Tu; G-proteins; Guanine Nucleotide Exchange Factor (GEF); Protein Synthesis; Ternary Complex; Transfer RNA (tRNA); Translation Regulation.
Figures



References
-
- Bourne H. R., Sanders D. A., McCormick F. (1991) The GTPase superfamily conserved structure and molecular mechanism. Nature 349, 117–127 - PubMed
-
- Vetter I. R., Wittinghofer A. (2001) The guanine nucleotide-binding switch in three dimensions. Science 294, 1299–1304 - PubMed
-
- Bos J. L., Rehmann H., Wittinghofer A. (2007) GEFs and GAPs: critical elements in the control of small G proteins. Cell 129, 865–877 - PubMed
-
- Wittinghofer A., Vetter I. R. (2011) Structure-function relationships of the G domain, a canonical switch motif. Annu. Rev. Biochem. 80, 943–971 - PubMed
-
- Thomposon R. C., Dix D. B. (1982) Accuracy of protein biosynthesis. A kinetic study of the reaction of poly(U)-programmed ribosomes with a leucyl-tRNA2-elongation factor Tu-GTP complex. J. Biol. Chem. 257, 6677–6682 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
