

A synthetic gene drive system for local, reversible modification and suppression of insect populations
- PMID: 23541732
- PMCID: PMC8459379
- DOI: 10.1016/j.cub.2013.02.059
A synthetic gene drive system for local, reversible modification and suppression of insect populations
Abstract
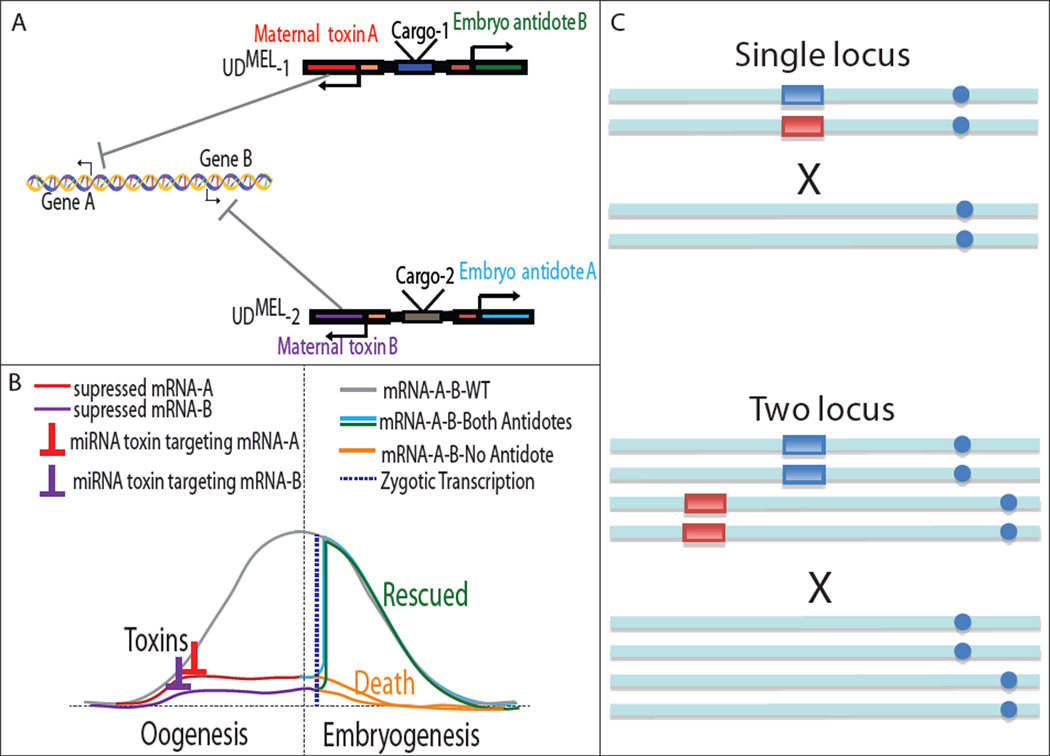
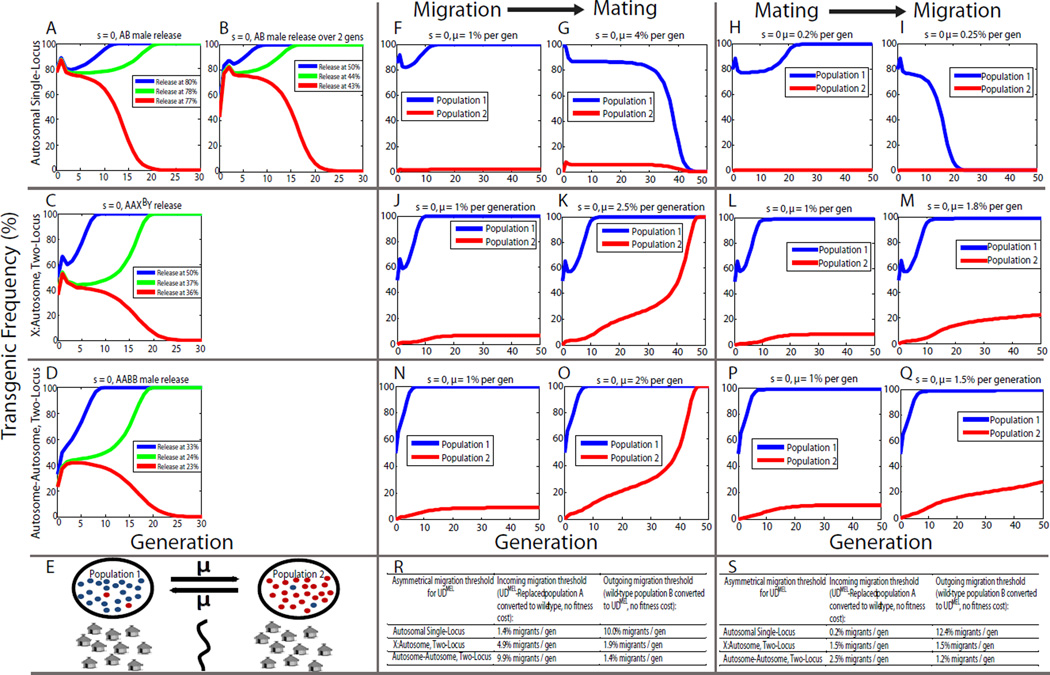
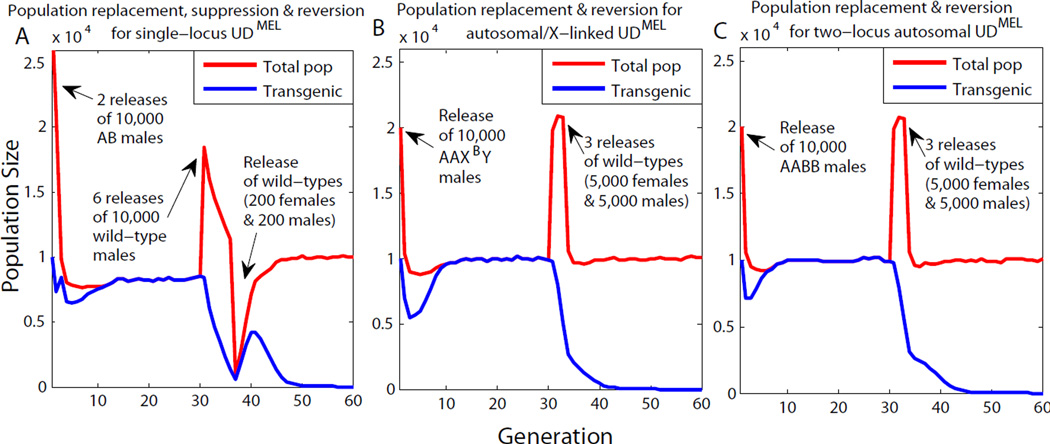
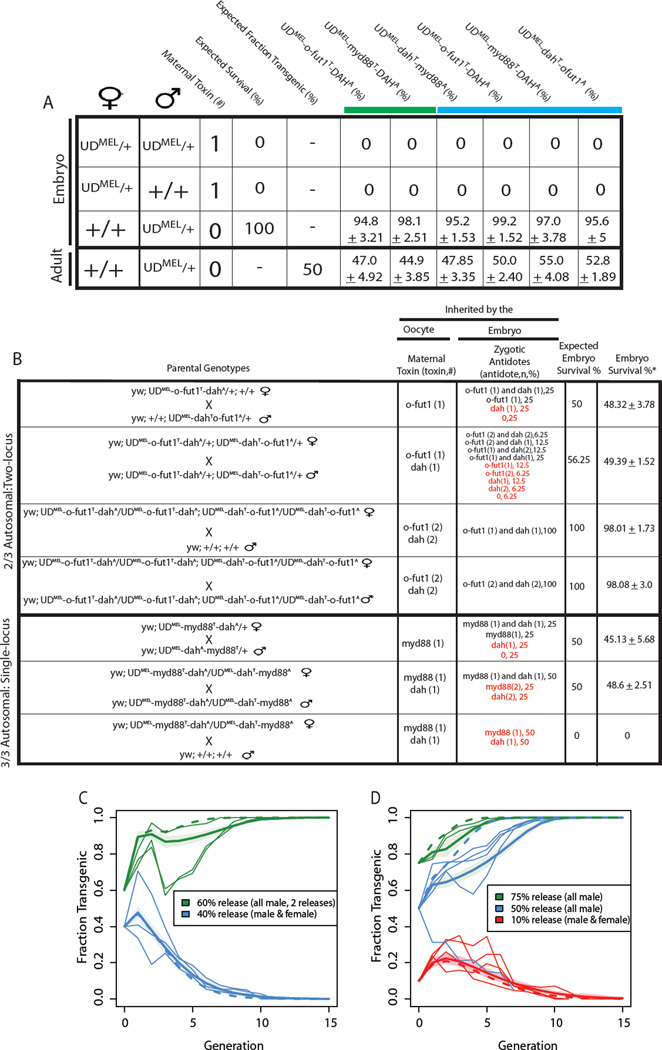
Replacement of wild insect populations with genetically modified individuals unable to transmit disease provides a self-perpetuating method of disease prevention but requires a gene drive mechanism to spread these traits to high frequency. Drive mechanisms requiring that transgenes exceed a threshold frequency in order to spread are attractive because they bring about local but not global replacement, and transgenes can be eliminated through dilution of the population with wild-type individuals. These features are likely to be important in many social and regulatory contexts. Here we describe the first creation of a synthetic threshold-dependent gene drive system, designated maternal-effect lethal underdominance (UD(MEL)), in which two maternally expressed toxins, located on separate chromosomes, are each linked with a zygotic antidote able to rescue maternal-effect lethality of the other toxin. We demonstrate threshold-dependent replacement in single- and two-locus configurations in Drosophila. Models suggest that transgene spread can often be limited to local environments. They also show that in a population in which single-locus UD(MEL) has been carried out, repeated release of wild-type males can result in population suppression, a novel method of genetic population manipulation.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Insect biotechnology: controllable replacement of disease vectors.Curr Biol. 2013 May 20;23(10):R453-6. doi: 10.1016/j.cub.2013.03.058. Curr Biol. 2013. PMID: 23701690
References
-
- Braig HR, Yan G. The spread of genetic constructs in natural insect populations. In: Letourneau DK, Burrows BE, editors. Genetically Engineered Organisms: Assessing Environmental and Human Health Effects. CRC Press; 2001. pp. 251–314.
-
- Sinkins SP, Gould F. Gene drive systems for insect disease vectors. Nat Rev Genet. 2006;7:427–435. - PubMed
-
- Altrock PM, Traulsen A, Reeves RG, Reed FA. Using underdominance to bi-stably transform local populations. J Theor Biol. 2010;267:62–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
