

Steric complementarity in the decoding center is important for tRNA selection by the ribosome
- PMID: 23542008
- PMCID: PMC3744617
- DOI: 10.1016/j.jmb.2013.02.038
Steric complementarity in the decoding center is important for tRNA selection by the ribosome
Abstract
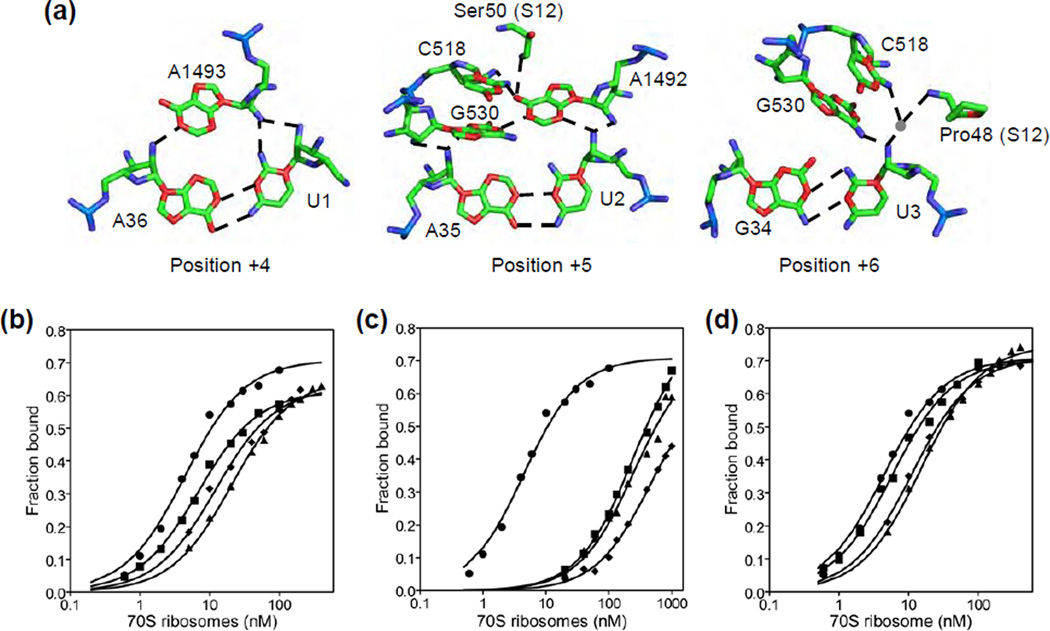
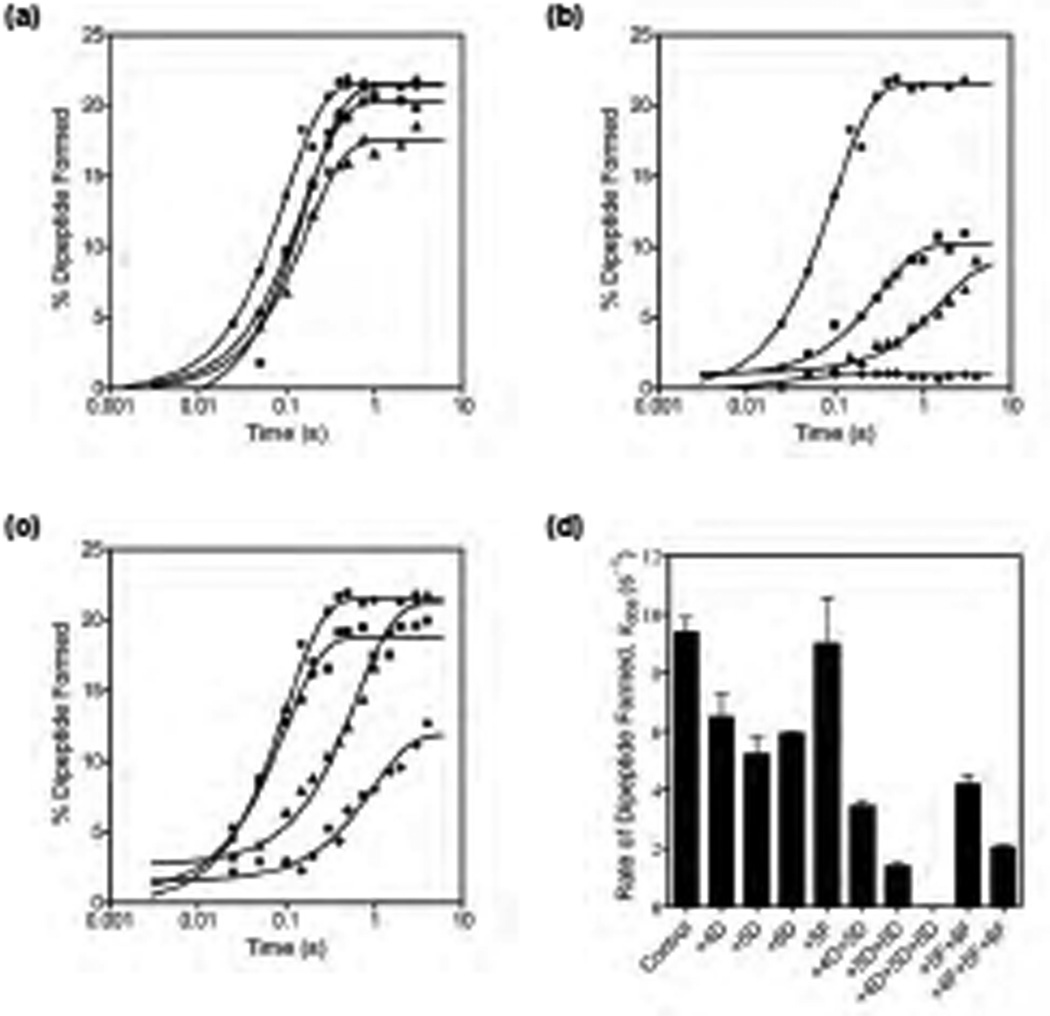
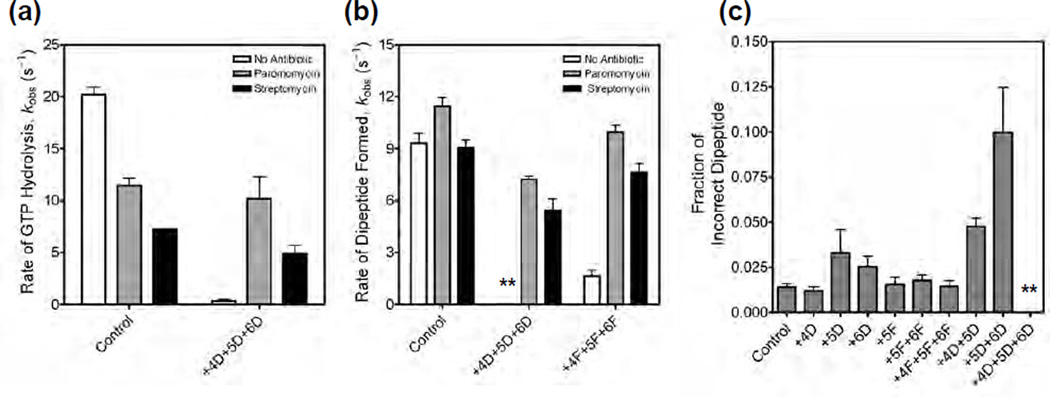
Accurate tRNA selection by the ribosome is essential for the synthesis of functional proteins. Previous structural studies indicated that the ribosome distinguishes between cognate and near-cognate tRNAs by monitoring the geometry of the codon-anticodon helix in the decoding center using the universally conserved 16S ribosomal RNA bases G530, A1492 and A1493. These bases form hydrogen bonds with the 2'-hydroxyl groups of the codon-anticodon helix, which are expected to be disrupted with a near-cognate codon-anticodon helix. However, a recent structural study showed that G530, A1492 and A1493 form hydrogen bonds in a manner identical with that of both cognate and near-cognate codon-anticodon helices. To understand how the ribosome discriminates between cognate and near-cognate tRNAs, we made 2'-deoxynucleotide and 2'-fluoro substituted mRNAs, which disrupt the hydrogen bonds between the A site codon and G530, A1492 and A1493. Our results show that multiple 2'-deoxynucleotide substitutions in the mRNA substantially inhibit tRNA selection, whereas multiple 2'-fluoro substitutions in the mRNA have only modest effects on tRNA selection. Furthermore, the miscoding antibiotics paromomycin and streptomycin rescue the defects in tRNA selection with the multiple 2'-deoxynucleotide substituted mRNA. These results suggest that steric complementarity in the decoding center is more important than the hydrogen bonds between the A site codon and G530, A1492 and A1493 for tRNA selection.
Keywords: EF-Tu; GTP hydrolysis; elongation factor Tu; kinetics; peptide bond; protein synthesis; rRNA; ribosomal RNA.
Copyright © 2013 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures

Similar articles
-
Selective perturbation of G530 of 16 S rRNA by translational miscoding agents and a streptomycin-dependence mutation in protein S12.J Mol Biol. 1994 Jan 7;235(1):156-72. doi: 10.1016/s0022-2836(05)80023-3. J Mol Biol. 1994. PMID: 8289238
-
A new understanding of the decoding principle on the ribosome.Nature. 2012 Mar 21;484(7393):256-9. doi: 10.1038/nature10913. Nature. 2012. PMID: 22437501
-
Recognition of cognate transfer RNA by the 30S ribosomal subunit.Science. 2001 May 4;292(5518):897-902. doi: 10.1126/science.1060612. Science. 2001. PMID: 11340196
-
Structural dynamics of ribosomal RNA during decoding on the ribosome.Biochimie. 2002 Aug;84(8):745-54. doi: 10.1016/s0300-9084(02)01409-8. Biochimie. 2002. PMID: 12457562 Review.
-
Insights into the decoding mechanism from recent ribosome structures.Trends Biochem Sci. 2003 May;28(5):259-66. doi: 10.1016/S0968-0004(03)00066-5. Trends Biochem Sci. 2003. PMID: 12765838 Review.
Cited by
-
Ribosome dynamics during decoding.Philos Trans R Soc Lond B Biol Sci. 2017 Mar 19;372(1716):20160182. doi: 10.1098/rstb.2016.0182. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28138068 Free PMC article. Review.
-
The parable of the caveman and the Ferrari: protein synthesis and the RNA world.Philos Trans R Soc Lond B Biol Sci. 2017 Mar 19;372(1716):20160187. doi: 10.1098/rstb.2016.0187. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28138073 Free PMC article. Review.
-
Elongation Factor Tu Switch I Element is a Gate for Aminoacyl-tRNA Selection.J Mol Biol. 2020 Apr 17;432(9):3064-3077. doi: 10.1016/j.jmb.2020.01.038. Epub 2020 Feb 13. J Mol Biol. 2020. PMID: 32061931 Free PMC article.
-
Genetic code degeneracy is established by the decoding center of the ribosome.Nucleic Acids Res. 2022 Apr 22;50(7):4113-4126. doi: 10.1093/nar/gkac171. Nucleic Acids Res. 2022. PMID: 35325219 Free PMC article.
-
EF-G catalyzes tRNA translocation by disrupting interactions between decoding center and codon-anticodon duplex.Nat Struct Mol Biol. 2014 Sep;21(9):817-24. doi: 10.1038/nsmb.2869. Epub 2014 Aug 10. Nat Struct Mol Biol. 2014. PMID: 25108354
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
