

Increased axonal bouton dynamics in the aging mouse cortex
- PMID: 23542382
- PMCID: PMC3631669
- DOI: 10.1073/pnas.1218731110
Increased axonal bouton dynamics in the aging mouse cortex
Abstract
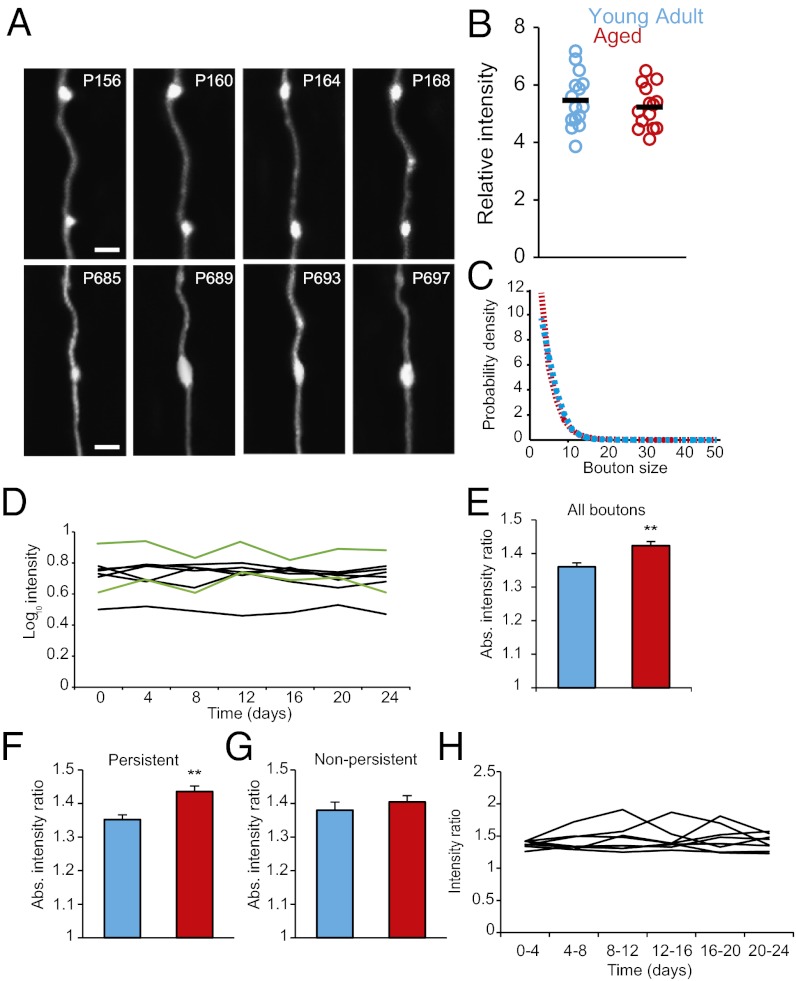
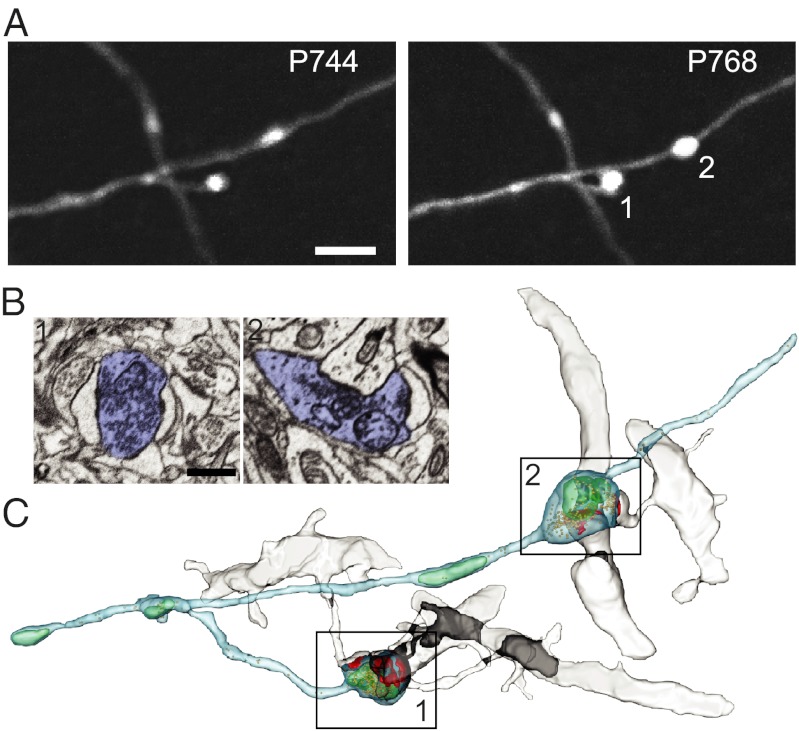
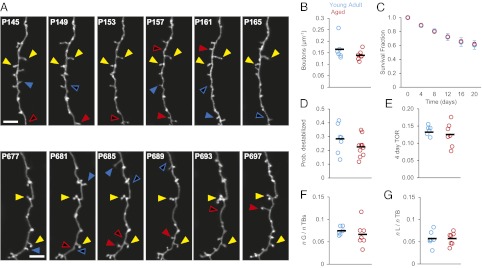
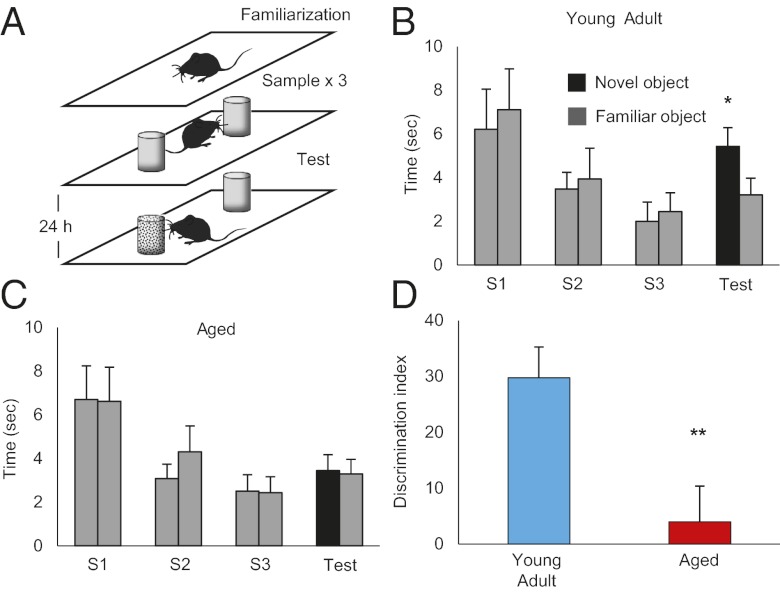
Aging is a major risk factor for many neurological diseases and is associated with mild cognitive decline. Previous studies suggest that aging is accompanied by reduced synapse number and synaptic plasticity in specific brain regions. However, most studies, to date, used either postmortem or ex vivo preparations and lacked key in vivo evidence. Thus, whether neuronal arbors and synaptic structures remain dynamic in the intact aged brain and whether specific synaptic deficits arise during aging remains unknown. Here we used in vivo two-photon imaging and a unique analysis method to rigorously measure and track the size and location of axonal boutons in aged mice. Unexpectedly, the aged cortex shows circuit-specific increased rates of axonal bouton formation, elimination, and destabilization. Compared with the young adult brain, large (i.e., strong) boutons show 10-fold higher rates of destabilization and 20-fold higher turnover in the aged cortex. Size fluctuations of persistent boutons, believed to encode long-term memories, also are larger in the aged brain, whereas bouton size and density are not affected. Our data uncover a striking and unexpected increase in axonal bouton dynamics in the aged cortex. The increased turnover and destabilization rates of large boutons indicate that learning and memory deficits in the aged brain arise not through an inability to form new synapses but rather through decreased synaptic tenacity. Overall our study suggests that increased synaptic structural dynamics in specific cortical circuits may be a mechanism for age-related cognitive decline.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
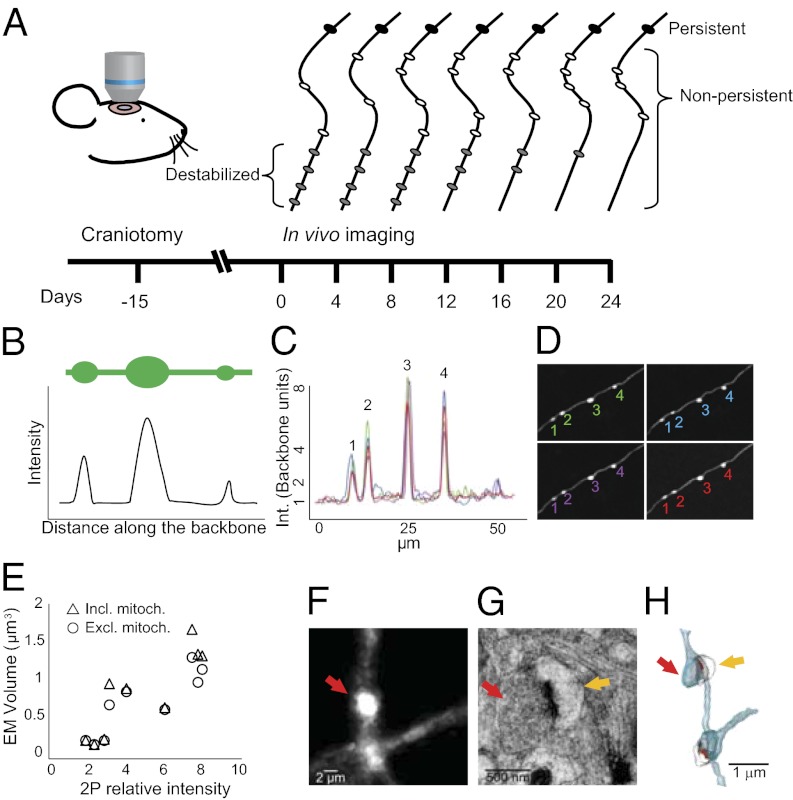
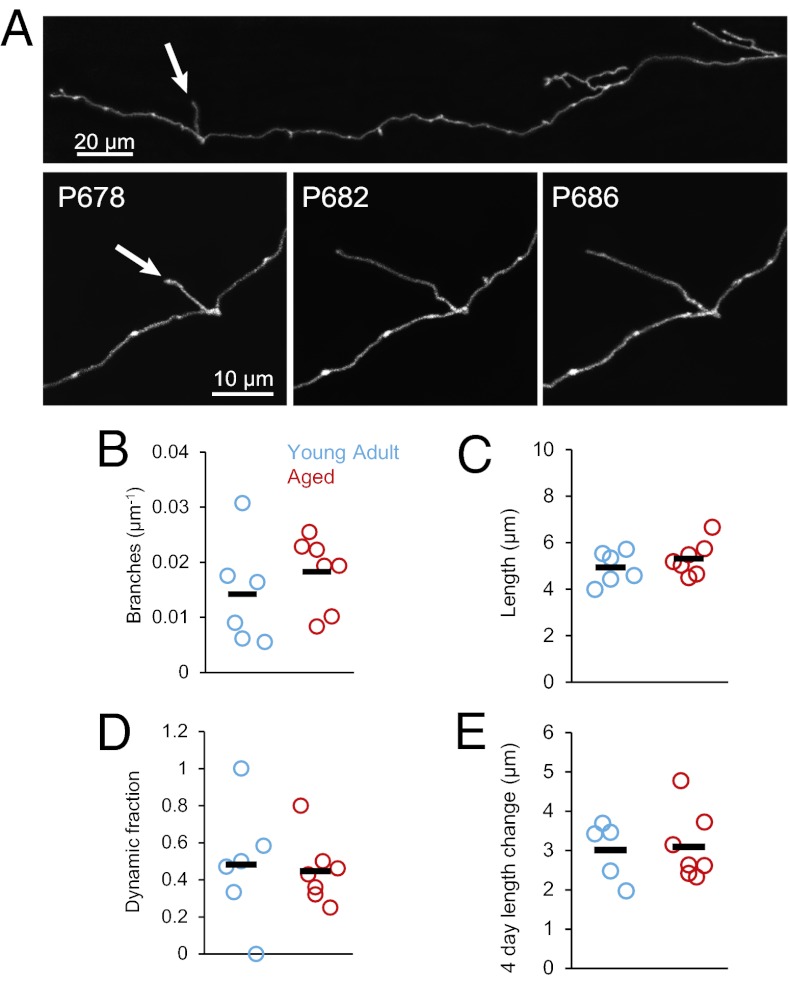
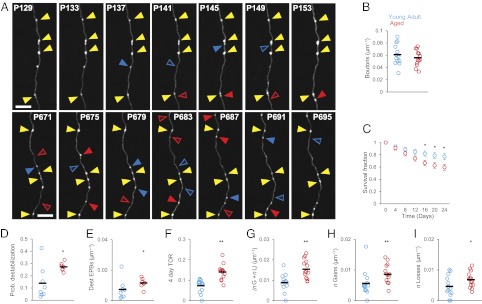
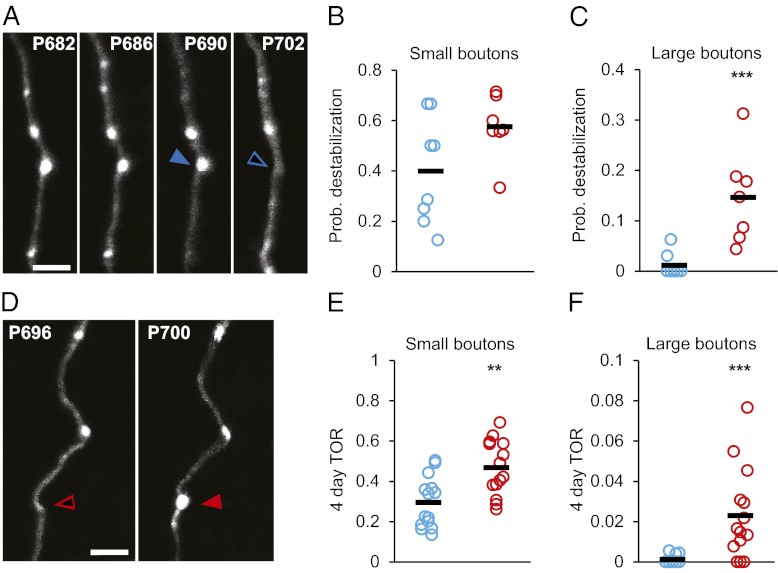
References
-
- Burke SN, Barnes CA. Neural plasticity in the ageing brain. Nat Rev Neurosci. 2006;7(1):30–40. - PubMed
-
- Geinisman Y, deToledo-Morrell L, Morrell F, Persina IS, Rossi M. Age-related loss of axospinous synapses formed by two afferent systems in the rat dentate gyrus as revealed by the unbiased stereological dissector technique. Hippocampus. 1992;2(4):437–444. - PubMed
-
- Foster TC, Barnes CA, Rao G, McNaughton BL. Increase in perforant path quantal size in aged F-344 rats. Neurobiol Aging. 1991;12(5):441–448. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
