

Mutations in STAMBP, encoding a deubiquitinating enzyme, cause microcephaly-capillary malformation syndrome
- PMID: 23542699
- PMCID: PMC4000253
- DOI: 10.1038/ng.2602
Mutations in STAMBP, encoding a deubiquitinating enzyme, cause microcephaly-capillary malformation syndrome
Abstract
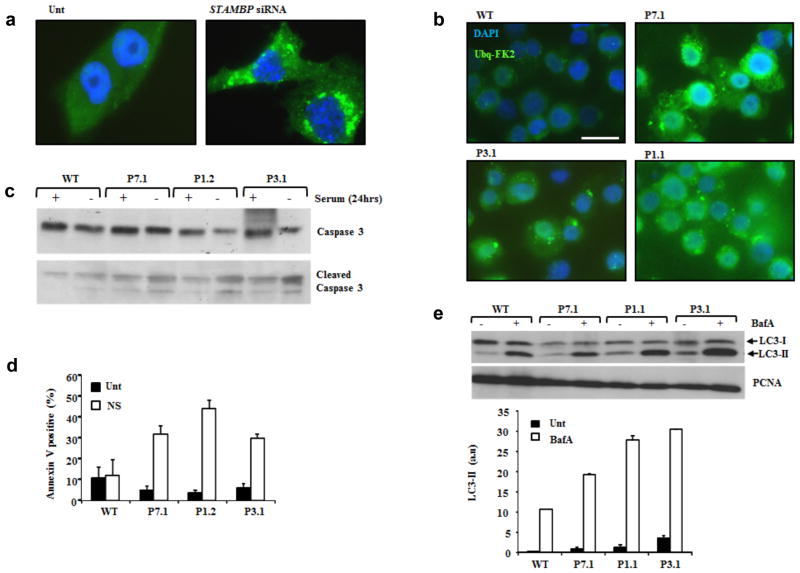
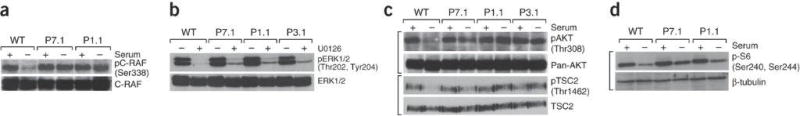
Microcephaly-capillary malformation (MIC-CAP) syndrome is characterized by severe microcephaly with progressive cortical atrophy, intractable epilepsy, profound developmental delay and multiple small capillary malformations on the skin. We used whole-exome sequencing of five patients with MIC-CAP syndrome and identified recessive mutations in STAMBP, a gene encoding the deubiquitinating (DUB) isopeptidase STAMBP (STAM-binding protein, also known as AMSH, associated molecule with the SH3 domain of STAM) that has a key role in cell surface receptor-mediated endocytosis and sorting. Patient cell lines showed reduced STAMBP expression associated with accumulation of ubiquitin-conjugated protein aggregates, elevated apoptosis and insensitive activation of the RAS-MAPK and PI3K-AKT-mTOR pathways. The latter cellular phenotype is notable considering the established connection between these pathways and their association with vascular and capillary malformations. Furthermore, our findings of a congenital human disorder caused by a defective DUB protein that functions in endocytosis implicates ubiquitin-conjugate aggregation and elevated apoptosis as factors potentially influencing the progressive neuronal loss underlying MIC-CAP syndrome.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



References
-
- Carter MT, et al. A new syndrome with multiple capillary malformations, intractable seizures, and brain and limb anomalies. Am J Med Genet A. 2011;155:301–306. - PubMed
-
- Isidor B, Barbarot S, Bénéteau C, Le Caignec C, David A. Multiple capillary skin malformations, epilepsy, microcephaly, mental retardation, hypoplasia of the distal phalanges: Report of a new case and further delineation of a new syndrome. Am J Med Genet A. 2011;155:1458–1460. - PubMed
-
- Carter MT, Boycot KM. Microcephaly–capillary malformation syndrome: A story of rapid emergence of a new recognizable entity. Am J Med Genet A. 2011;155:2078–2079. - PubMed
-
- Jacobs AH, Walton RG. The incidence of birthmarks in the neonate. Pediatrics. 1976;58:218–222. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
