

Inhibition of PIKfyve by YM-201636 dysregulates autophagy and leads to apoptosis-independent neuronal cell death
- PMID: 23544129
- PMCID: PMC3609765
- DOI: 10.1371/journal.pone.0060152
Inhibition of PIKfyve by YM-201636 dysregulates autophagy and leads to apoptosis-independent neuronal cell death
Abstract
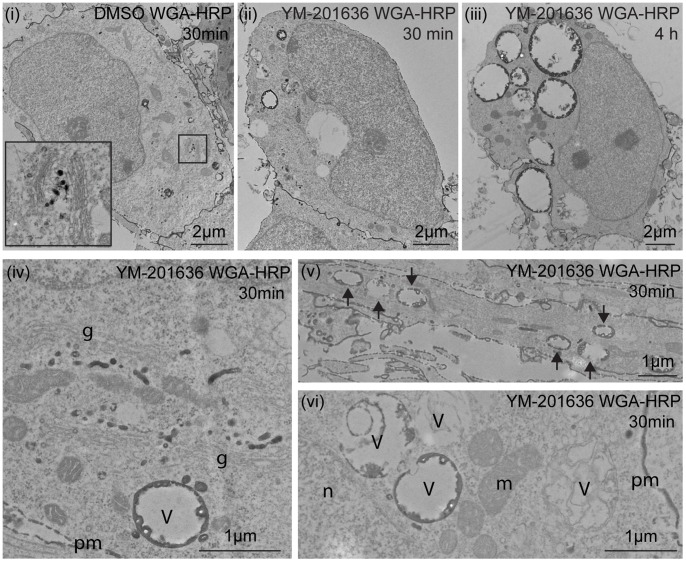
The lipid phosphatidylinositol 3,5-bisphosphate (PtdIns(3,5)P 2), synthesised by PIKfyve, regulates a number of intracellular membrane trafficking pathways. Genetic alteration of the PIKfyve complex, leading to even a mild reduction in PtdIns(3,5)P 2, results in marked neurodegeneration via an uncharacterised mechanism. In the present study we have shown that selectively inhibiting PIKfyve activity, using YM-201636, significantly reduces the survival of primary mouse hippocampal neurons in culture. YM-201636 treatment promoted vacuolation of endolysosomal membranes followed by apoptosis-independent cell death. Many vacuoles contained intravacuolar membranes and inclusions reminiscent of autolysosomes. Accordingly, YM-201636 treatment increased the level of the autophagosomal marker protein LC3-II, an effect that was potentiated by inhibition of lysosomal proteases, suggesting that alterations in autophagy could be a contributing factor to neuronal cell death.
Conflict of interest statement
Figures






References
-
- Sbrissa D, Ikonomov OC, Fu Z, Ijuin T, Gruenberg J, et al. (2007) Core protein machinery for mammalian phosphatidylinositol 3,5-bisphosphate synthesis and turnover that regulates the progression of endosomal transport. Novel Sac phosphatase joins the ArPIKfyve-PIKfyve complex. J Biol Chem 282: 23878–23891. - PubMed
-
- Ikonomov OC, Sbrissa D, Shisheva A (2001) Mammalian cell morphology and endocytic membrane homeostasis require enzymatically active phosphoinositide 5-kinase PIKfyve. J Biol Chem 276: 26141–26147. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
