

The Brucella abortus general stress response system regulates chronic mammalian infection and is controlled by phosphorylation and proteolysis
- PMID: 23546883
- PMCID: PMC3650426
- DOI: 10.1074/jbc.M113.459305
The Brucella abortus general stress response system regulates chronic mammalian infection and is controlled by phosphorylation and proteolysis
Abstract
Background: Virulence of pathogenic bacteria is often determined by their ability to adapt to stress.
Results: The Brucella abortus general stress response (GSR) system is required for chronic mammalian infection and is regulated by phosphorylation and proteolysis.
Conclusion: The B. abortus GSR signaling pathway has multiple layers of post-translational control and is a determinant of chronic infection.
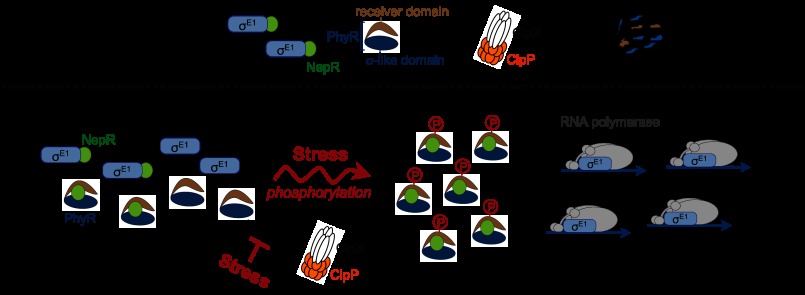
Significance: This study provides new, molecular level insight into chronic Brucella infection. Brucella spp. are adept at establishing a chronic infection in mammals. We demonstrate that core components of the α-proteobacterial general stress response (GSR) system, PhyR and σ(E1), are required for Brucella abortus stress survival in vitro and maintenance of chronic murine infection in vivo. ΔphyR and ΔrpoE1 null mutants exhibit decreased survival under acute oxidative and acid stress but are not defective in infection of primary murine macrophages or in initial colonization of BALB/c mouse spleens. However, ΔphyR and ΔrpoE1 mutants are attenuated in spleens beginning 1 month postinfection. Thus, the B. abortus GSR system is dispensable for colonization but is required to maintain chronic infection. A genome-scale analysis of the B. abortus GSR regulon identified stress response genes previously linked to virulence and genes that affect immunomodulatory components of the cell envelope. These data support a model in which the GSR system affects both stress survival and the interface between B. abortus and the host immune system. We further demonstrate that PhyR proteolysis is a unique feature of GSR control in B. abortus. Proteolysis of PhyR provides a mechanism to avoid spurious PhyR protein interactions that inappropriately activate GSR-dependent transcription. We conclude that the B. abortus GSR system regulates acute stress adaptation and long term survival within a mammalian host and that PhyR proteolysis is a novel regulatory feature in B. abortus that ensures proper control of GSR transcription.
Keywords: Bacterial Pathogenesis; Bacterial Signal Transduction; Bacterial Transcription; Brucella; General Stress Response; Infectious Diseases; PhyR; Stress Response.
Figures






Similar articles
-
The Brucella abortus virulence regulator, LovhK, is a sensor kinase in the general stress response signalling pathway.Mol Microbiol. 2014 Nov;94(4):913-25. doi: 10.1111/mmi.12809. Epub 2014 Oct 19. Mol Microbiol. 2014. PMID: 25257300 Free PMC article.
-
WrpA Is an Atypical Flavodoxin Family Protein under Regulatory Control of the Brucella abortus General Stress Response System.J Bacteriol. 2016 Mar 31;198(8):1281-93. doi: 10.1128/JB.00982-15. Print 2016 Apr. J Bacteriol. 2016. PMID: 26858101 Free PMC article.
-
Brucella abortus ΔrpoE1 confers protective immunity against wild type challenge in a mouse model of brucellosis.Vaccine. 2016 Sep 30;34(42):5073-5081. doi: 10.1016/j.vaccine.2016.08.076. Epub 2016 Aug 31. Vaccine. 2016. PMID: 27591954 Free PMC article.
-
Brucella stationary-phase gene expression and virulence.Annu Rev Microbiol. 2003;57:57-76. doi: 10.1146/annurev.micro.57.030502.090803. Epub 2003 May 1. Annu Rev Microbiol. 2003. PMID: 12730323 Review.
-
Brucella abortus Cell Cycle and Infection Are Coordinated.Trends Microbiol. 2015 Dec;23(12):812-821. doi: 10.1016/j.tim.2015.09.007. Epub 2015 Oct 20. Trends Microbiol. 2015. PMID: 26497941 Free PMC article. Review.
Cited by
-
A Carbonic Anhydrase Pseudogene Sensitizes Select Brucella Lineages to Low CO2 Tension.J Bacteriol. 2019 Oct 21;201(22):e00509-19. doi: 10.1128/JB.00509-19. Print 2019 Nov 15. J Bacteriol. 2019. PMID: 31481543 Free PMC article.
-
The functional differences between paralogous regulators define the control of the general stress response in Sphingopyxis granuli TFA.Environ Microbiol. 2022 Apr;24(4):1918-1931. doi: 10.1111/1462-2920.15907. Epub 2022 Jan 27. Environ Microbiol. 2022. PMID: 35049124 Free PMC article.
-
The Brucella Cell Envelope.Annu Rev Microbiol. 2023 Sep 15;77:233-253. doi: 10.1146/annurev-micro-032521-013159. Epub 2023 Apr 27. Annu Rev Microbiol. 2023. PMID: 37104660 Free PMC article. Review.
-
Brucellosis: Bacteriology, pathogenesis, epidemiology and role of the metallophores in virulence: a review.Front Cell Infect Microbiol. 2025 Jul 8;15:1621230. doi: 10.3389/fcimb.2025.1621230. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 40697821 Free PMC article. Review.
-
An Extracytoplasmic Function Sigma/Anti-Sigma Factor System Regulates Hypochlorous Acid Resistance and Impacts Expression of the Type IV Secretion System in Brucella melitensis.J Bacteriol. 2021 May 20;203(12):e0012721. doi: 10.1128/JB.00127-21. Epub 2021 Apr 5. J Bacteriol. 2021. PMID: 33820796 Free PMC article.
References
-
- Pappas G., Papadimitriou P., Akritidis N., Christou L., Tsianos E. V. (2006) The new global map of human brucellosis. Lancet Infect. Dis. 6, 91–99 - PubMed
-
- Roop R. M., 2nd, Bellaire B. H., Valderas M. W., Cardelli J. A. (2004) Adaptation of the brucellae to their intracellular niche. Mol. Microbiol. 52, 621–630 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
