

Review
doi: 10.1083/jcb.201301061.
Aneuploidy in health, disease, and aging
Affiliations
- PMID: 23547028
- PMCID: PMC3613689
- DOI: 10.1083/jcb.201301061
Item in Clipboard
Review
Aneuploidy in health, disease, and aging
J Cell Biol.
.
Abstract
Aneuploidy, an aberrant number of chromosomes, has been recognized as a feature of human malignancies for over a century, but compelling evidence for causality was largely lacking until mouse models for chromosome number instability were used. These in vivo studies have not only uncovered important new insights into the extremely complex aneuploidy-cancer relationship but also into the molecular mechanisms underlying proper and aberrant chromosome segregation. A series of diverse mouse models for the mitotic checkpoint protein BubR1 has provided evidence for a provocative novel link between aneuploidization and the development of age-related pathologies.
Figures
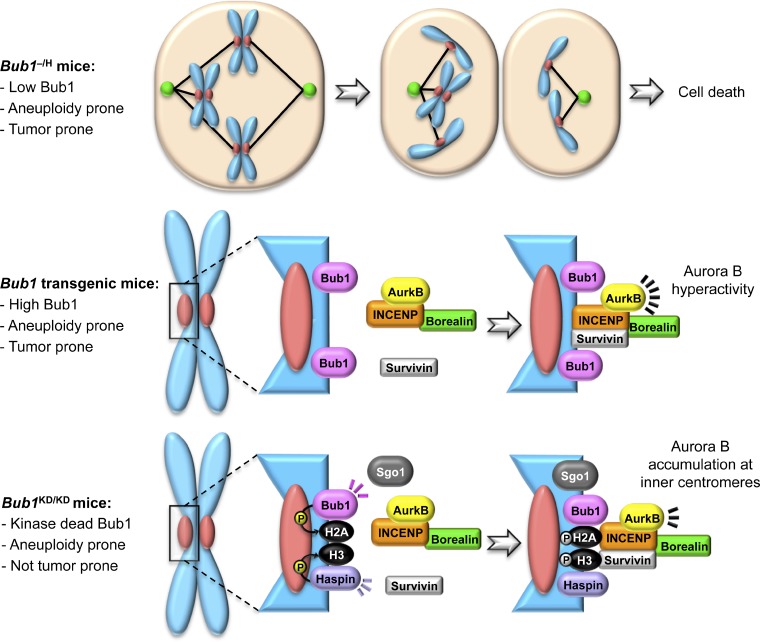
New mechanistic insights from various Bub1 mouse models. Bidirectional deviations from normal Bub1 levels and inactivation of Bub1 enzymatic activity universally cause aneuploid cells to accumulate in mice. Analysis of the underlying mechanisms of chromosome missegregation in each mouse model has provided important new insights into the multifaceted nature of this mitotic regulator. For instance, in addition to confirming that Bub1 plays a critical role in kinetochore assembly and mitotic checkpoint activity, studies of Bub1 hypomorphic mice (top) revealed that Bub1 acts as a crucial trigger to induce cell death after chromosome missegregation (Jeganathan et al., 2007). On the other hand, transgenic mice that overexpress Bub1 (middle) revealed the novel concept of Aurora B hyperactivity and linked it to chromosome missegregation (Ricke et al., 2011). Unlike Bub1 hypomorphic or transgenic mice, mice lacking Bub1 kinase activity (bottom) harbor significant aneuploidy without a predisposition to cancer. In this model, it was revealed that accumulation of Aurora B at inner centromeric regions is mediated by Bub1-mediated histone H2A phosphorylation at T121 in a shugoshin-independent manner (Ricke et al., 2012). P, phosphorylation.
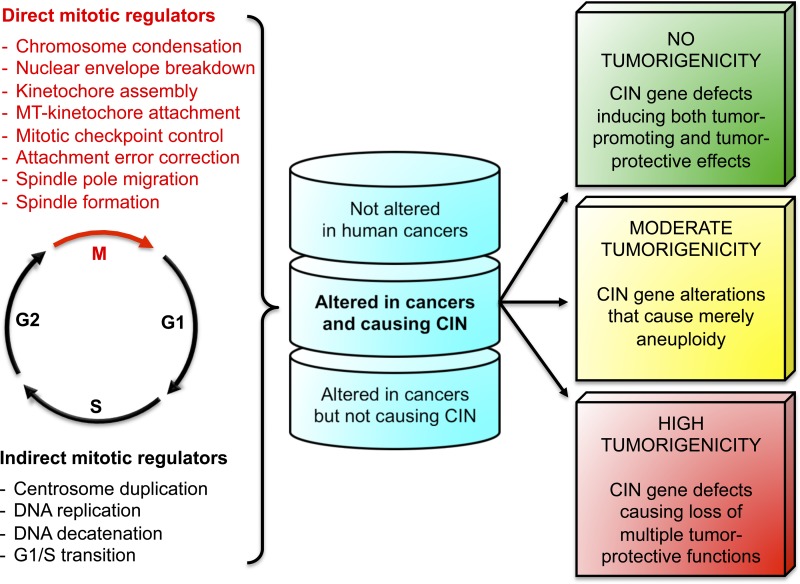
A hierarchy-based view of the aneuploidy–cancer relationship. Hundreds of genes are thought to directly or indirectly influence chromosomal stability. Whereas the functions of direct mitotic regulators are limited to mitosis, cellular processes acting outside of mitosis that influence segregation accuracy are defined as indirect mitotic regulators. Genetic modification of CIN genes in mice suggests that certain gene alterations are more cancer relevant than others, leading us to propose a hierarchal CIN gene model. This model not only takes into consideration aneuploidy rates but also other cancer-critical functions CIN genes might possess and the frequency with which a particular CIN gene alteration occurs in human cancers. We recognize three classes of CIN gene alterations within both the direct and indirect mitotic regulators: CIN genes that are not or rarely found altered in human cancers, CIN gene alterations found in human cancers but not causing aneuploidy, and genes that are altered and act causally to promote chromosome missegregation. Animal modeling studies predict three potential cancer-related outcomes for the latter class of CIN gene alterations: (1) no tumorigenicity (or inhibiting tumor development), (2) moderate tumorigenicity, and (3) high tumorigenicity. The latter CIN gene defects presumably are multifaceted and act through numerous tumor-protective functions, whereas those with only moderate tumorigenicity perhaps act solely in the generation of aneuploid cells. Alternatively, CIN genes with counteracting tumor-promoting and tumor-protecting functions may have no tumorigenicity predisposition. One example for this final type is BubR1, through which deregulation induces both aneuploidization and cellular senescence.
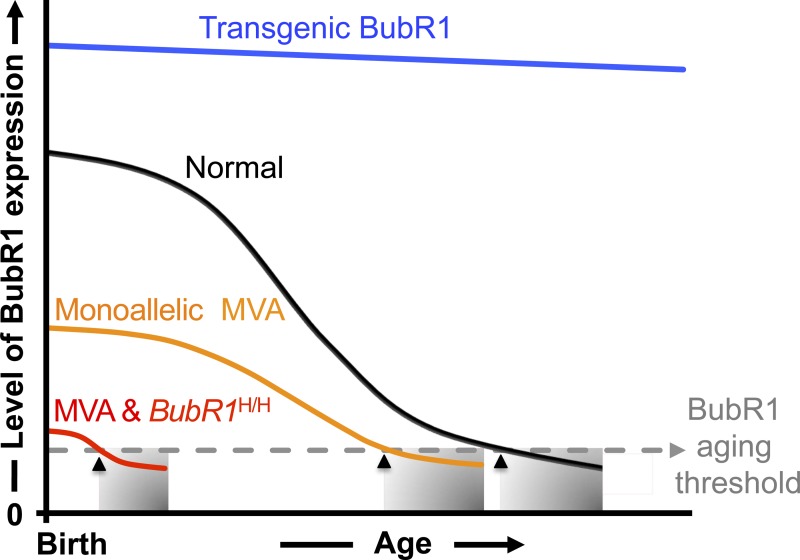
A minimum threshold level of BubR1 delays age-related pathologies. Several lines of evidence provide a link between BubR1 level and time of age-related pathology onset. During natural aging (black line), BubR1 levels decline in various tissues (Baker et al., 2004, 2013). Mutant mice with very low levels of BubR1 at birth prematurely develop age-related phenotypes with a shortened lifespan (red line). Similarly, many patients with mosaic variegated aneuploidy (MVA) harbor mutations within BubR1 that diminish steady-state BubR1 protein levels (Suijkerbuijk et al., 2010). Even the presence of a single MVA allele negatively impacts health- and lifespan in mice (Wijshake et al., 2012), implying that carriers of an MVA mutation may reach the critical BubR1 threshold level earlier than normal individuals (orange line). Another line of evidence that the amount of BubR1 impacts health has been revealed through artificially elevating BubR1 to extend both health- and lifespan (blue line; Baker et al., 2004, 2013). Together, these results suggest that a threshold level of BubR1 (gray dashed line) is required to prevent onset (black arrows) of tissue deterioration (shaded regions).
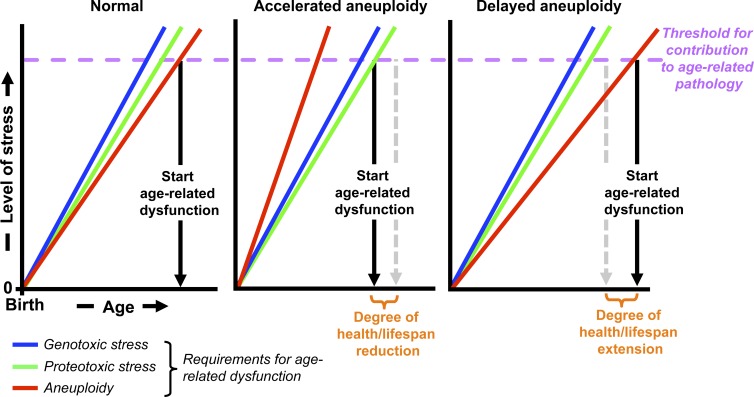
Hypothetical models for how aneuploidy might impact health- and lifespan. Working model for how aneuploidy rates might modulate age-related tissue dysfunction. The model takes into consideration the widely held view that tissue aging is driven, at least in part, by diverse cellular stresses causing progressive accumulation of various types of macromolecular damage/stress (Finkel et al., 2007). For simplicity, only three kinds of stresses have been included in the model: aneuploidy, proteotoxic, and genotoxic stress. Accelerated aneuploidy caused by CIN in and of itself is not sufficient to initiate age-related deterioration but may expedite aging by reducing the time needed to reach the cumulative stress threshold. On the other hand, delayed aneuploidization, such as through increased BubR1, may attenuate the aneuploidy stress component and reduce the combined stress sum, thereby extending healthy lifespan. We note that accelerated aneuploidy may influence the threshold of other stresses to trigger pathology because aneuploidy itself can activate genotoxic and proteotoxic stress (Williams et al., 2008; Sheltzer et al., 2011; Tang et al., 2011). Thus, the rate of aneuploidy may alter the slope of other cellular stresses (not depicted).
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
