

Inactivation of the mTORC1-eukaryotic translation initiation factor 4E pathway alters stress granule formation
- PMID: 23547259
- PMCID: PMC3648080
- DOI: 10.1128/MCB.01517-12
Inactivation of the mTORC1-eukaryotic translation initiation factor 4E pathway alters stress granule formation
Abstract
Stress granules (SG) are cytoplasmic multimeric RNA bodies that form under stress conditions known to inhibit cap-dependent translation. SG contain translation initiation factors, RNA binding proteins, and signaling molecules. SG are known to inhibit apoptotic pathways, thus contributing to chemo- and radioresistance in tumor cells. However, whether stress granule formation involves oncogenic signaling pathways is currently unknown. Here, we report a novel role of the mTORC1-eukaryotic translation initiation factor 4E (eIF4E) pathway, a key regulator of cap-dependent translation initiation of oncogenic factors, in SG formation. mTORC1 specifically drives the eIF4E-mediated formation of SG through the phosphorylation of 4E-BP1, a key factor known to inhibit formation of the mTORC1-dependent eIF4E-eIF4GI interactions. Disrupting formation of SG by inactivation of mTOR with its specific inhibitor pp242 or by depletion of eIF4E or eIF4GI blocks the SG-associated antiapoptotic p21 pathway. Finally, pp242 sensitizes cancer cells to death in vitro and inhibits the growth of chemoresistant tumors in vivo. This work therefore highlights a novel role of the oncogenic mTORC1-eIF4E pathway, namely, the promotion of formation of antiapoptotic SG.
Figures
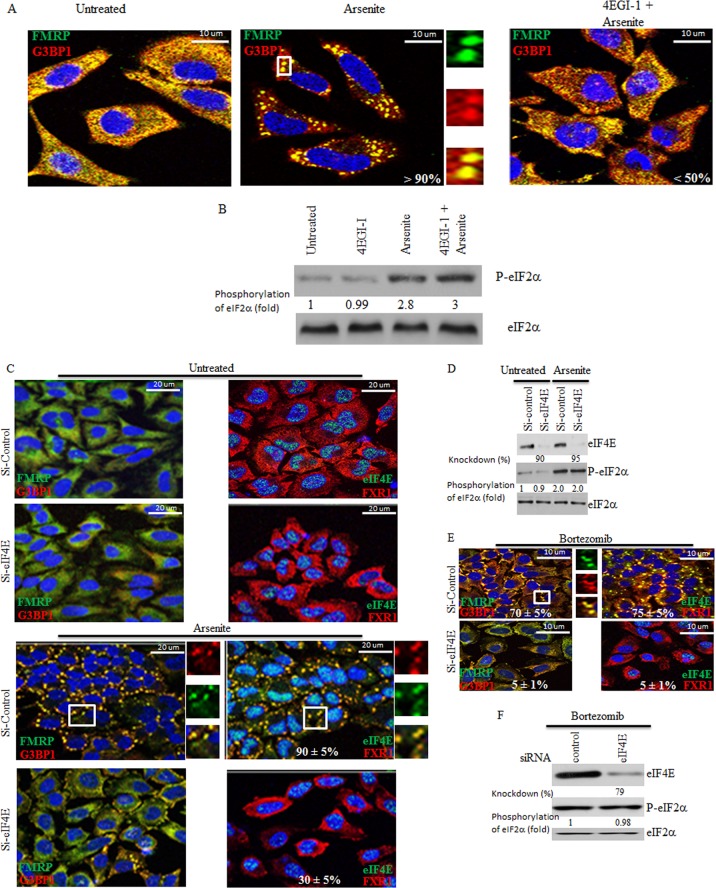
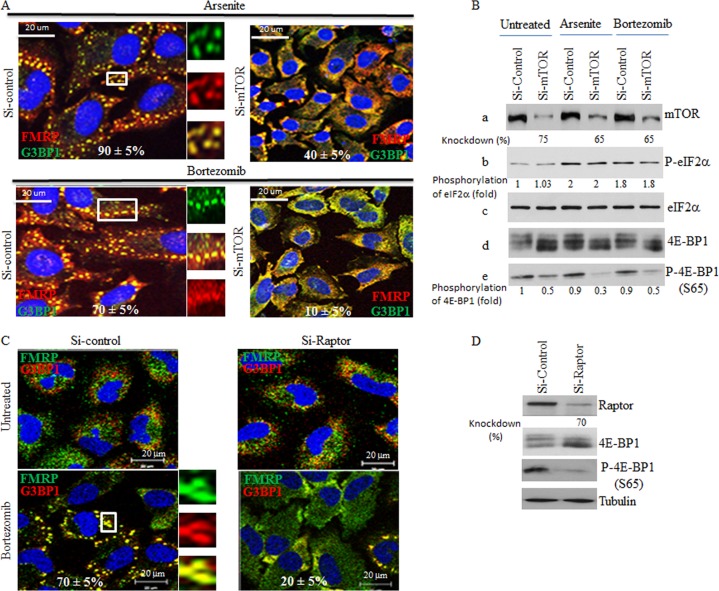
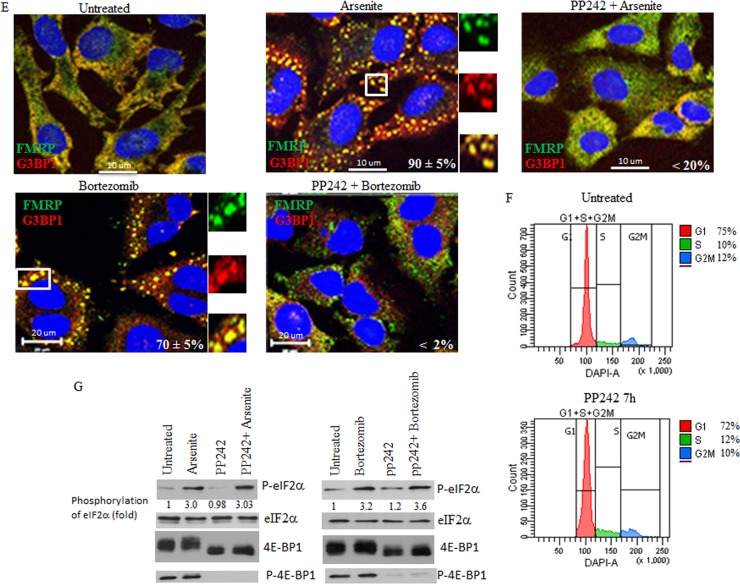
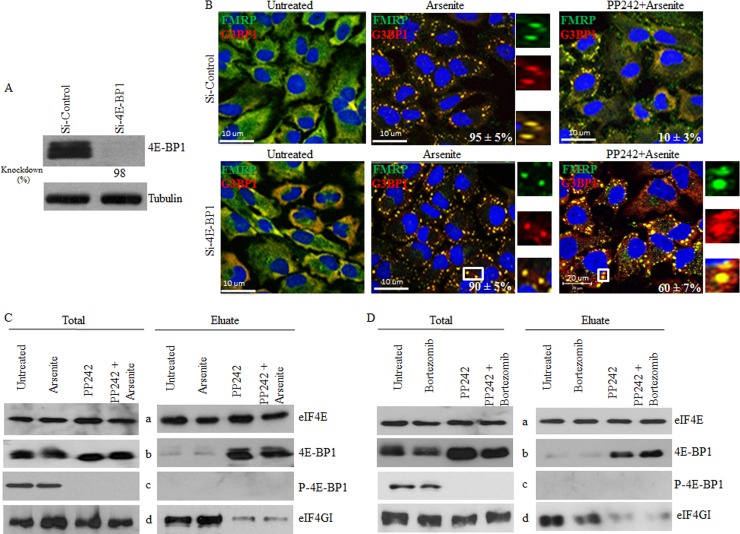
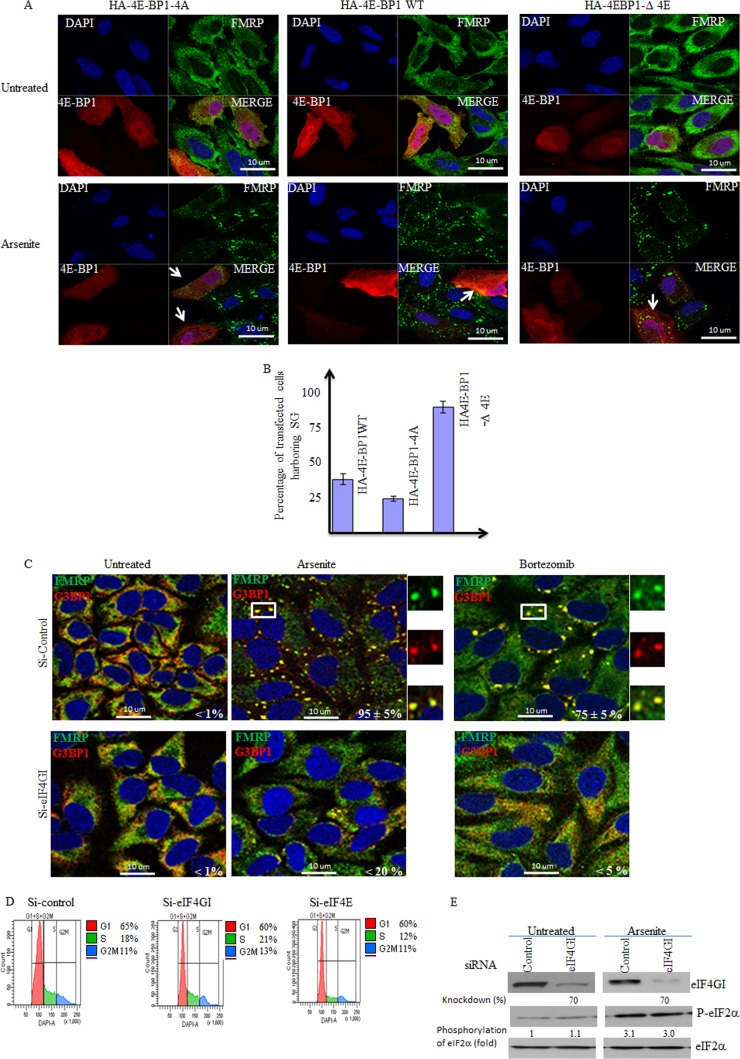
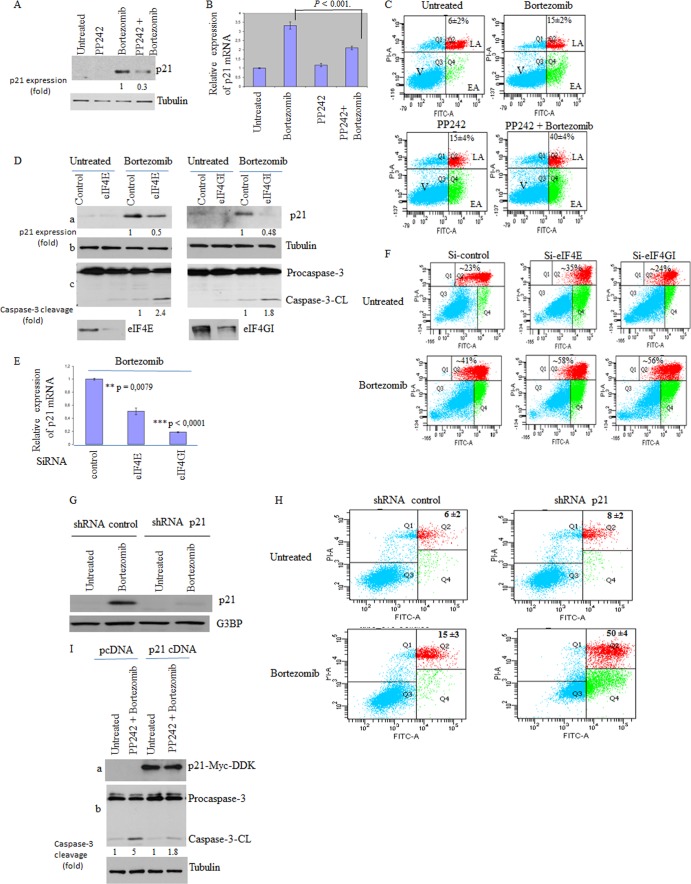
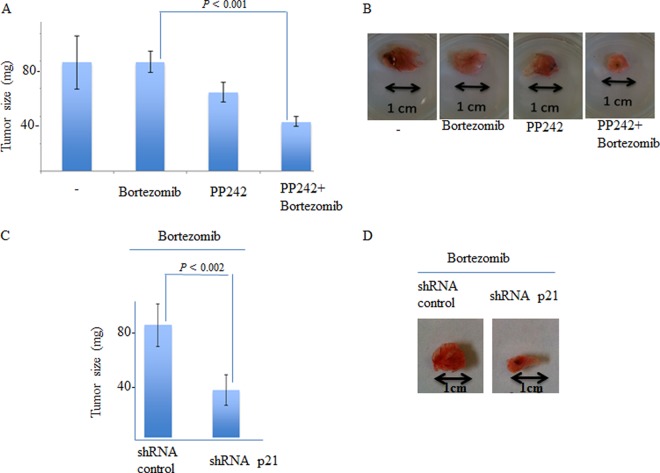
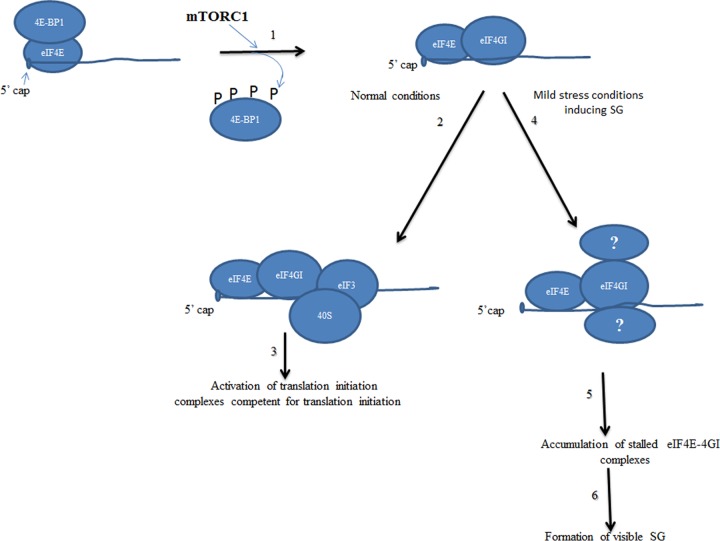
References
-
- Moeller BJ, Cao Y, Li CY, Dewhirst MW. 2004. Radiation activates HIF-1 to regulate vascular radiosensitivity in tumors: role of reoxygenation, free radicals, and stress granules. Cancer Cell 5:429–441 - PubMed
-
- Fournier MJ, Gareau C, Mazroui R. 2010. The chemotherapeutic agent bortezomib induces the formation of stress granules. Cancer Cell Int. 10:12 doi:10.1186/1475-2867-10-12 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous