

Kisspeptin and puberty in mammals
- PMID: 23550010
- PMCID: PMC3996677
- DOI: 10.1007/978-1-4614-6199-9_12
Kisspeptin and puberty in mammals
Abstract
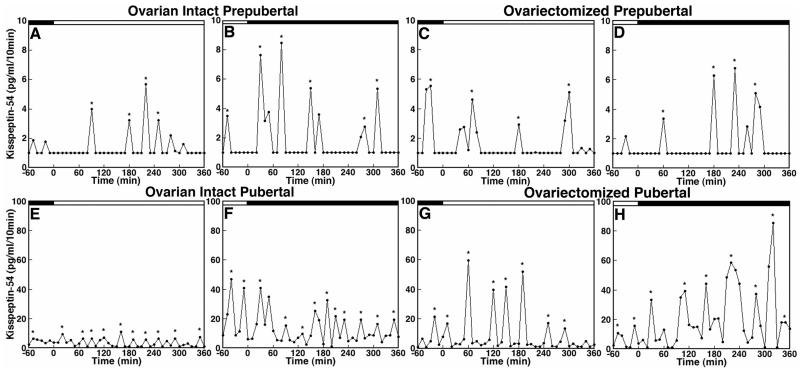
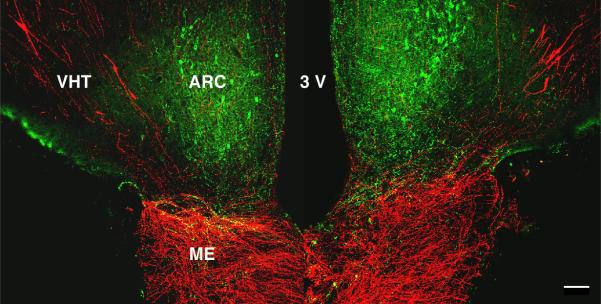
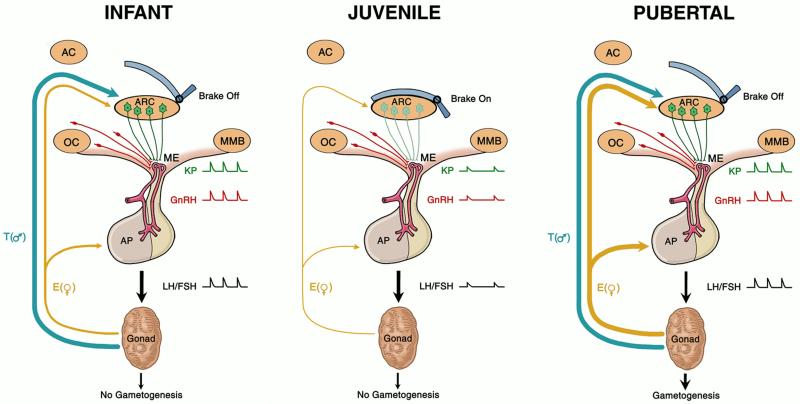
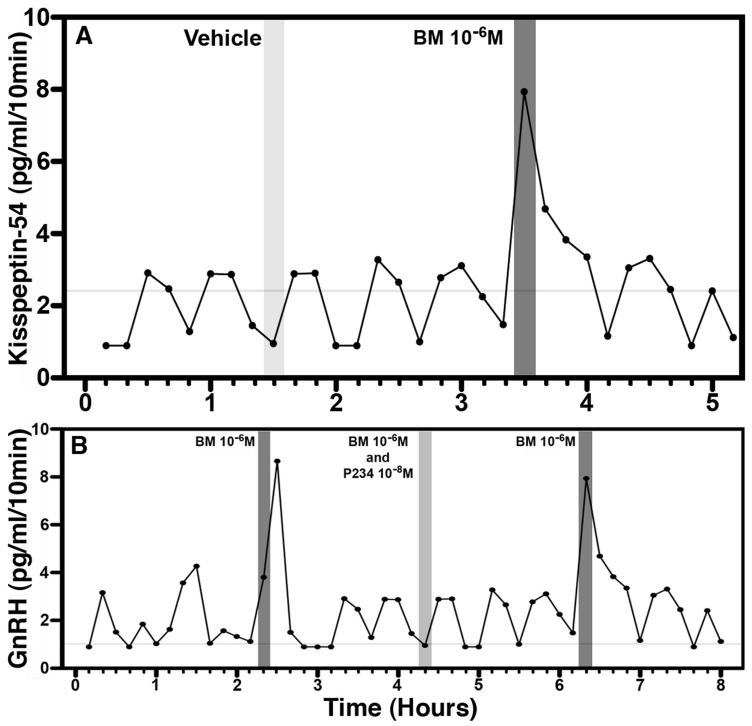
Since the discovery of the G-protein coupled receptor 54 (kisspeptin receptor) and its ligand, kisspeptin, our understanding of the neurobiological mechanisms that govern the pituitary-gonadal axis has evolved dramatically. In this chapter, we have reviewed progress regarding the relationship between kisspeptin and puberty, and have proposed a novel hypothesis for the role of kisspeptin signaling in the onset of this crucial developmental event. According to this hypothesis, although kisspeptin neurons in the arcuate nucleus (ARC) are critical for puberty, this is simply because these cells are an integral component of the hypothalamic GnRH pulse generating mechanism that drives intermittent release of the decapeptide, as an increase in GnRH is obligatory for the onset of puberty. In our model, ARC kisspeptin neurons play no "regulatory" role in controlling the timing of puberty. Rather, as a component of the neural network responsible for GnRH pulse generation, they subserve upstream regulatory mechanisms that are responsible for the timing of puberty.
Figures
References
-
- Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS, Jr, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O’Rahilly S, Carlton MB, Crowley WF, Jr, Aparicio SA, Colledge WH. The GPR54 gene as a regulator of puberty. N Engl J Med. 2003;349:1614–27. - PubMed
-
- Semple RK, Achermann JC, Ellery J, Farooqi IS, Karet FE, Stanhope RG, O’rahilly S, Aparicio SA. Novel missense mutations in g protein-coupled receptor 54 in a patient with hypogonadotropic hypogonadism. J Clin Endocrinol Metab. 2005;90:1849–55. - PubMed
-
- Cerrato F, Shagoury J, Kralickova M, Dwyer A, Falardeau J, Ozata M, Van Vliet G, Bouloux P, Hall JE, Hayes FJ, Pitteloud N, Martin KA, Welt C, Seminara SB. Coding sequence analysis of GnRHR and GPR54 in patients with congenital and adult onset forms of hypogonadotropic hypogonadism. Eur J Endocrinol. 2006;155:S3–S10. - PubMed
-
- Tenenbaum-Rakover Y, Commenges-Ducos M, Iovane A, Aumas C, Admoni O, de Roux N. Neuroendocrine phenotype analysis in five patients with isolated hypogonadotropic hypogonadism due to a L102P inactivating mutation of GPR54. J Clin Endocrinol Metab. 2007;92:1137–44. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
