

Review
doi: 10.1111/imr.12045.
MicroRNA function in NK-cell biology
Affiliations
- PMID: 23550637
- PMCID: PMC3621029
- DOI: 10.1111/imr.12045
Item in Clipboard
Review
MicroRNA function in NK-cell biology
Immunol Rev.
2013 May.
Abstract
The important role of microRNAs in directing immune responses has become increasingly clear. Here, we highlight discoveries uncovering the role of specific microRNAs in regulating the development and function of natural killer (NK) cells. Furthermore, we discuss the impact of NK cells on the entire immune system during global and specific microRNA ablation in the settings of inflammation, infection, and immune dysregulation.
© 2013 John Wiley & Sons A/S. Published by Blackwell Publishing Ltd.
Conflict of interest statement
The authors have no conflicts of interest to declare.
Figures
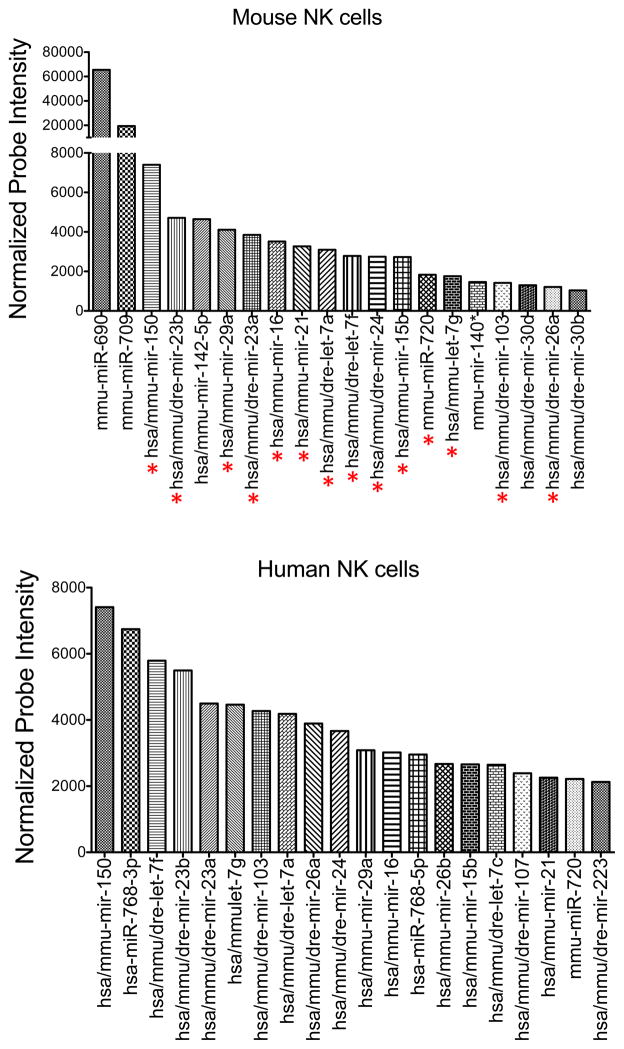
Total RNA was extracted from sorted mouse splenic NK cells (NK1.1+ TCRβ−) and human peripheral blood NK cells (CD56+CD3−). Expression of individual miRNAs was assessed by microarray. After normalization, 170 oligonucleotide probes gave mean fluorescence values above background. Shown are the normalized expression values (in arbitrary units) of the top 20 most highly expressed miRNAs in mouse (top) and human (bottom) NK cells. miRNAs common to both groups are indicated with a red asterisk below the top graph.
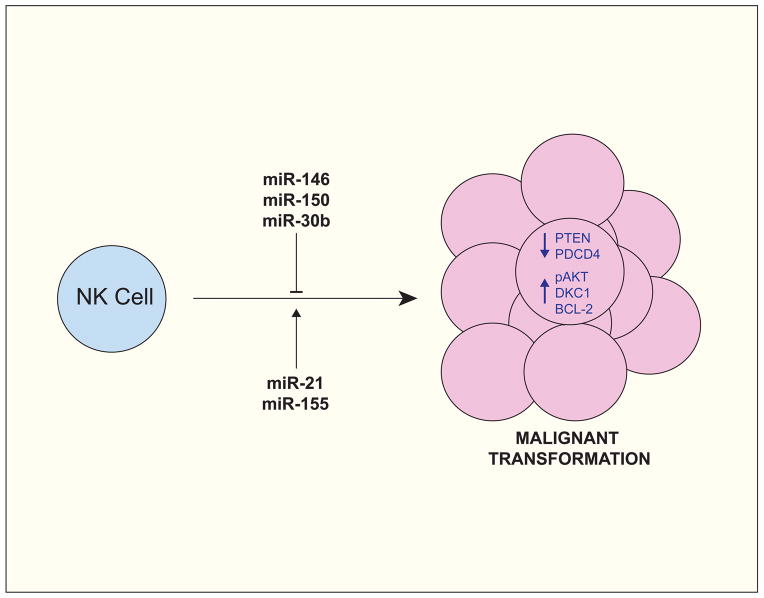
miR-150 and miR-21 are often over-expressed in Natural Killer/T cell Lymphomas (NKTLs) and have been shown to promote oncogenic transformation by repressing tumor suppressor factors (e.g. PTEN and PDCD4) and by promoting pro-proliferative pathways (e.g. the PI3K-AKT cascade). In contrast, miR-146, miR-150, and miR-30b, which are often under-expressed in NKTLs, have been attributed with tumor suppressor activity, owing in part to their ability to repress pro-proliferative and pro-survival factors, such as AKT2, DKC1, and, indirectly, BCL-2.
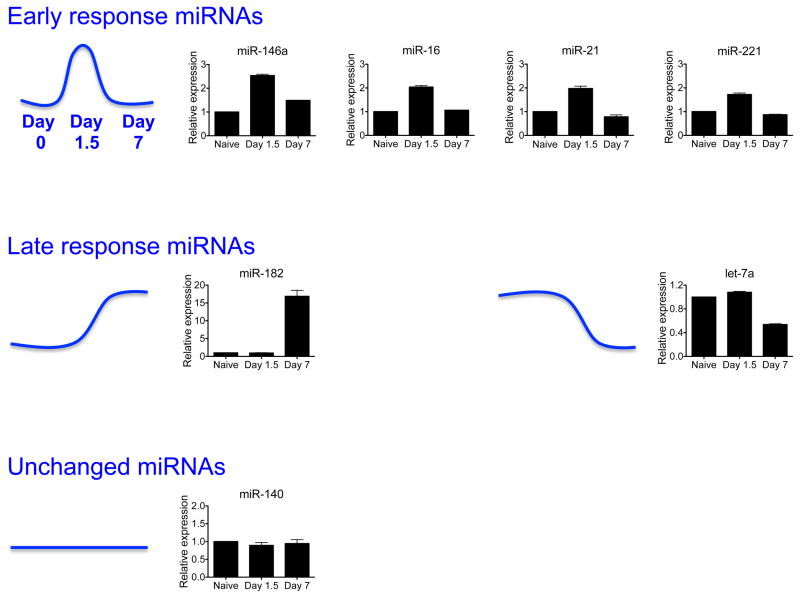
Expression of select miRNAs in splenic Ly49H+ NK cells sorted from animals infected with Smith strain MCMV, as assessed by qRT-PCR. Levels of mature miRNAs were normalized to sno-202. Data are shown as the fold change at day 1.5 and day 7 versus day 0 (naïve). miRNAs transiently upregulated on day 1.5 post-infection (i.e. Early response miRNAs) include miR-146a, miR-16, miR-21, and miR-221. miRNAs upregulated or downregulated on day 7 post-infection (i.e. Late response miRNAs) include miR-182 and let-7a. miR-140 levels remained constant on the assessed time points.
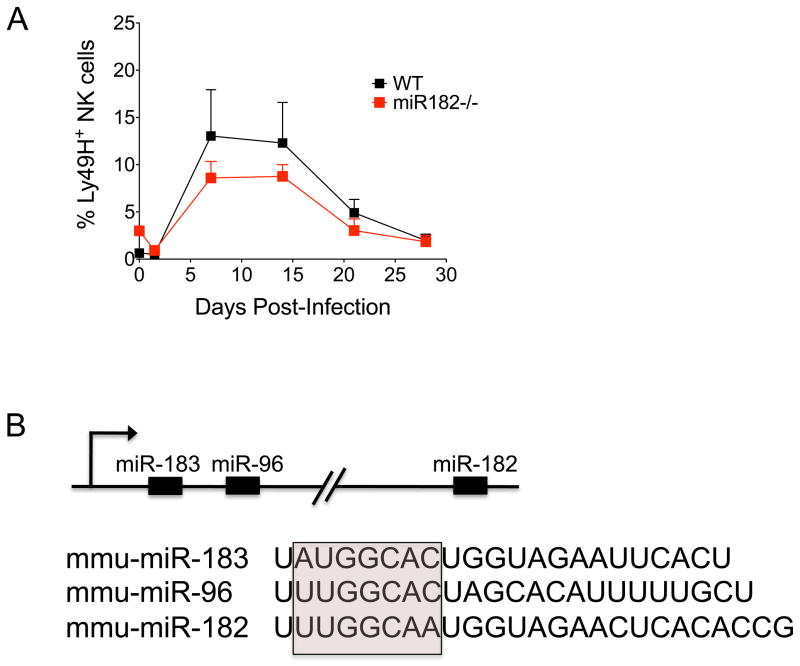
(A) Equal numbers of WT (CD45.1+) and miR-182−/− (CD45.2+) Ly49H+ NK cells were co-transferred into Ly49H-deficient hosts. Following infection with MCMV, the percentage of adoptively transferred WT (black) and miR-182−/−(red) Ly49H+ NK cells within the total NK cell population in blood was assessed at the indicated time points post-infection. Error bars show SEM (n = 3). Data are representative of three independent experiments. (B) Schematic representation of the miR-183 ~ 182 cluster. miR-183, miR-96, and miR-182 share sequence homology, particularly within the seed sequence used to bind target mRNAs (pink box).
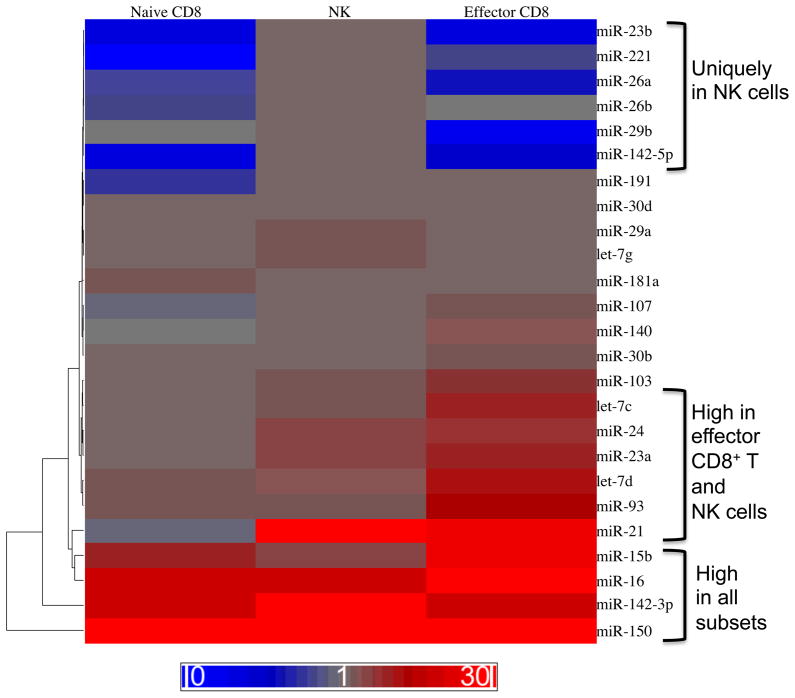
miRNAs expressed in resting NK cells as compared to resting (naive) CD8+ T cells or effector CD8+ T cells. NK1.1+ TCRβ− NK cells were sorted from C57BL/6 mice. Naive CD8+ T cells were sorted from CD45.1+CD45.2+ P14 mice bearing the DbGP33-specific TCR. Effector CD8+ T cells were sorted from chimeric mice generated by adoptive transfer of P14 T cells into C57BL/6 (CD45.2+) recipients that were infected intraperitoneally with 2 × 105 plaque forming units (PFU) of LCMV Armstrong. Levels of mature miRNAs were measured by qRT-PCR and normalized to sno202. Fold expression relative to sno-202 is depicted as a heat map. Three patterns
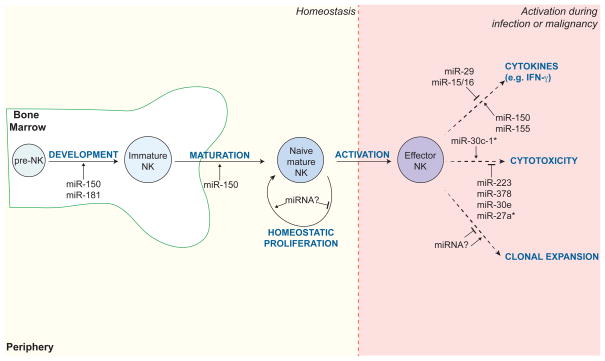
A summary of individual miRNAs known to regulate NK cell development and function at steady state and in the context of activation (e.g. during infection or malignancy). Examples include miR-150 and miR-181, which are required for NK cell development and maturation in the bone marrow; miR-150, miR-155, miR-29, and miR-15/16, which modulate cytokine production by activated NK cells; and miR-30c-1*, miR-223, miR-378, miR-30e, miR-27a*, which control expression of cytotoxic molecules, such as granzyme B and perforin.
References
-
- Orange JS. Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect. 2002;4:1545–1558. - PubMed
-
- Shanley JD. In vivo administration of monoclonal antibody to the NK 1. 1 antigen of natural killer cells: effect on acute murine cytomegalovirus infection. J Med Virol. 1990;30:58–60. - PubMed
-
- Ghiasi H, Cai S, Perng GC, Nesburn AB, Wechsler SL. The role of natural killer cells in protection of mice against death and corneal scarring following ocular HSV-1 infection. Antiviral Res. 2000;45:33–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
