

Parallel evolution of senescence in annual fishes in response to extrinsic mortality
- PMID: 23551990
- PMCID: PMC3623659
- DOI: 10.1186/1471-2148-13-77
Parallel evolution of senescence in annual fishes in response to extrinsic mortality
Abstract
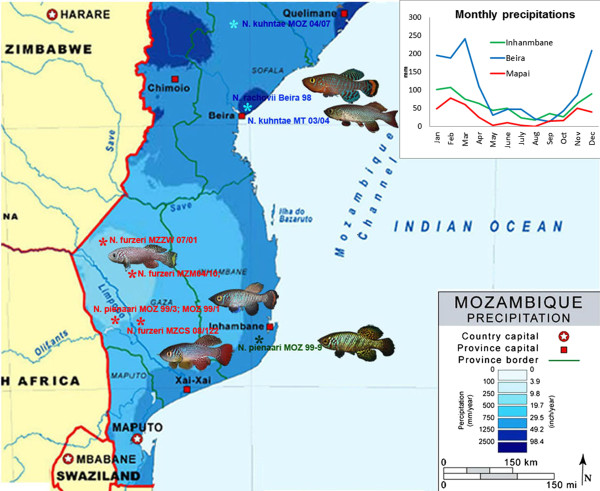
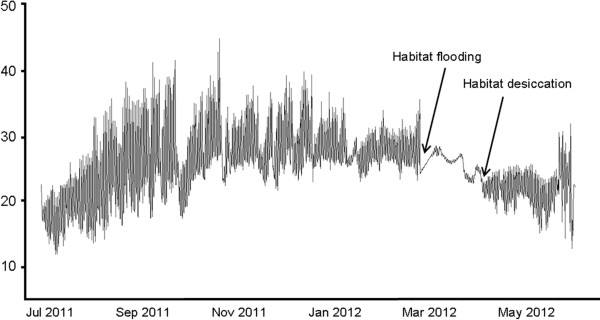
Background: Early evolutionary theories of aging predict that populations which experience low extrinsic mortality evolve a retarded onset of senescence. Experimental support for this theory in vertebrates is scarce, in part for the difficulty of quantifying extrinsic mortality and its condition- and density-dependent components that -when considered- can lead to predictions markedly different to those of the "classical" theories. Here, we study annual fish of the genus Nothobranchius whose maximum lifespan is dictated by the duration of the water bodies they inhabit. Different populations of annual fish do not experience different strengths of extrinsic mortality throughout their life span, but are subject to differential timing (and predictability) of a sudden habitat cessation. In this respect, our study allows testing how aging evolves in natural environments when populations vary in the prospect of survival, but condition-dependent survival has a limited effect. We use 10 Nothobranchius populations from seasonal pools that differ in their duration to test how this parameter affects longevity and aging in two independent clades of these annual fishes.
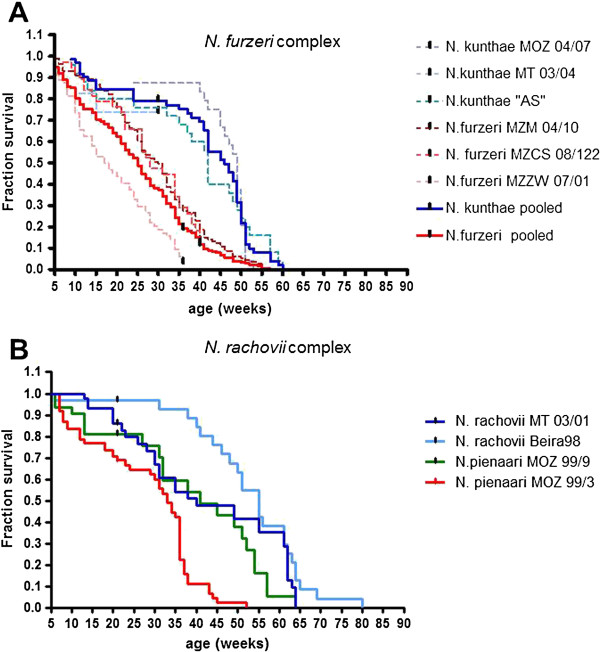
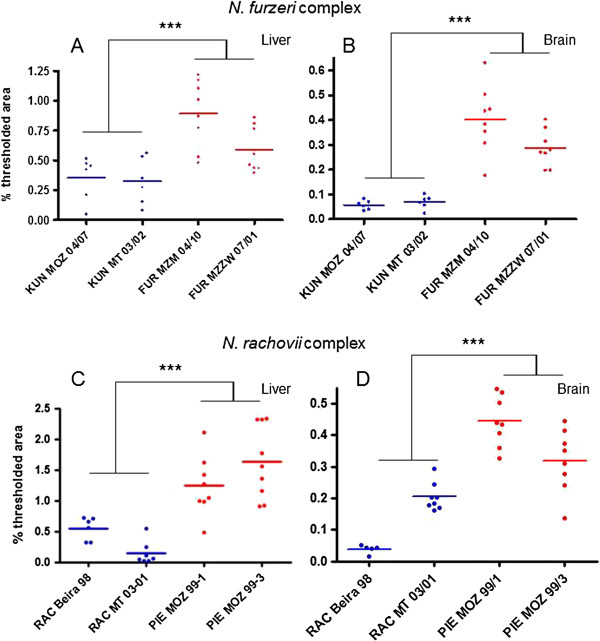
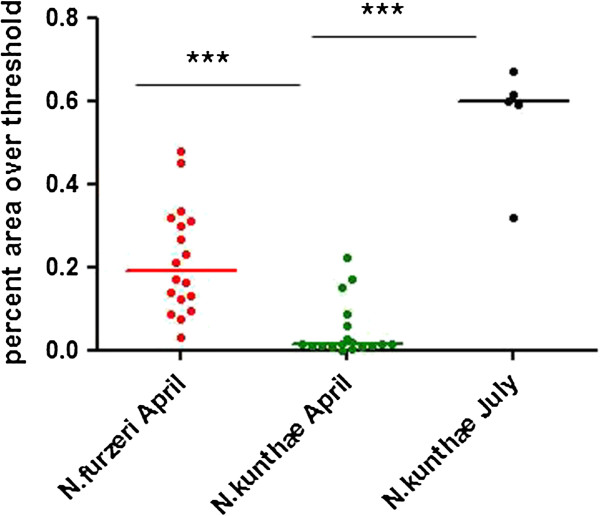
Results: We found that replicated populations from a dry region showed markedly shorter captive lifespan than populations from a humid region. Shorter lifespan correlated with accelerated accumulation of lipofuscin (an established age marker) in both clades. Analysis of wild individuals confirmed that fish from drier habitats accumulate lipofuscin faster also under natural conditions. This indicates faster physiological deterioration in shorter-lived populations.
Conclusions: Our data provide a strong quantitative example of how extrinsic mortality can shape evolution of senescence in a vertebrate clade. Nothobranchius is emerging as a genomic model species. The characterization of pairs of closely related species with different longevities should provide a powerful paradigm for the identification of genetic variations responsible for evolution of senescence in natural populations.
Figures
References
-
- Austad SN. Retarded senescence in an insular population of virginia opossums (didelphis-virginiana) J Zool. 1993;229:695–708. doi: 10.1111/j.1469-7998.1993.tb02665.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
