

The emergence of functional microcircuits in visual cortex
- PMID: 23552948
- PMCID: PMC4843961
- DOI: 10.1038/nature12015
The emergence of functional microcircuits in visual cortex
Abstract
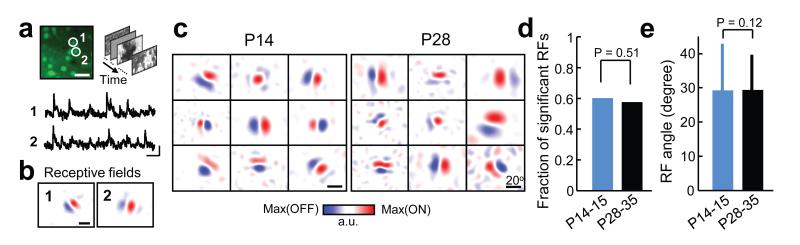
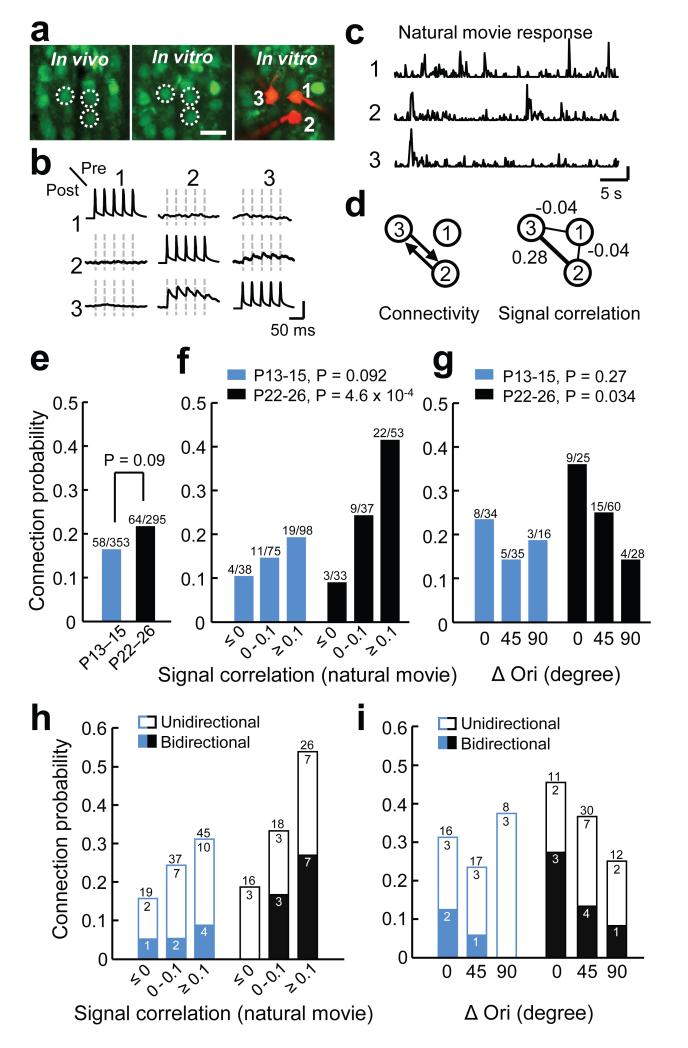
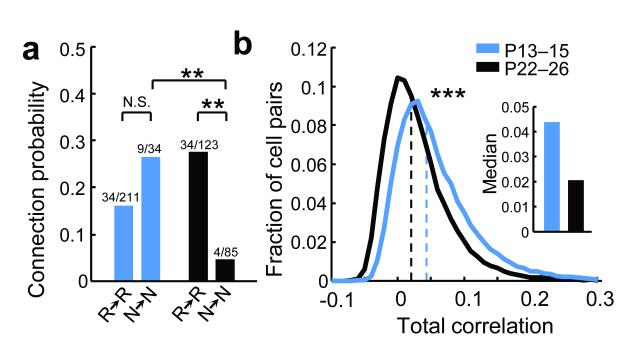
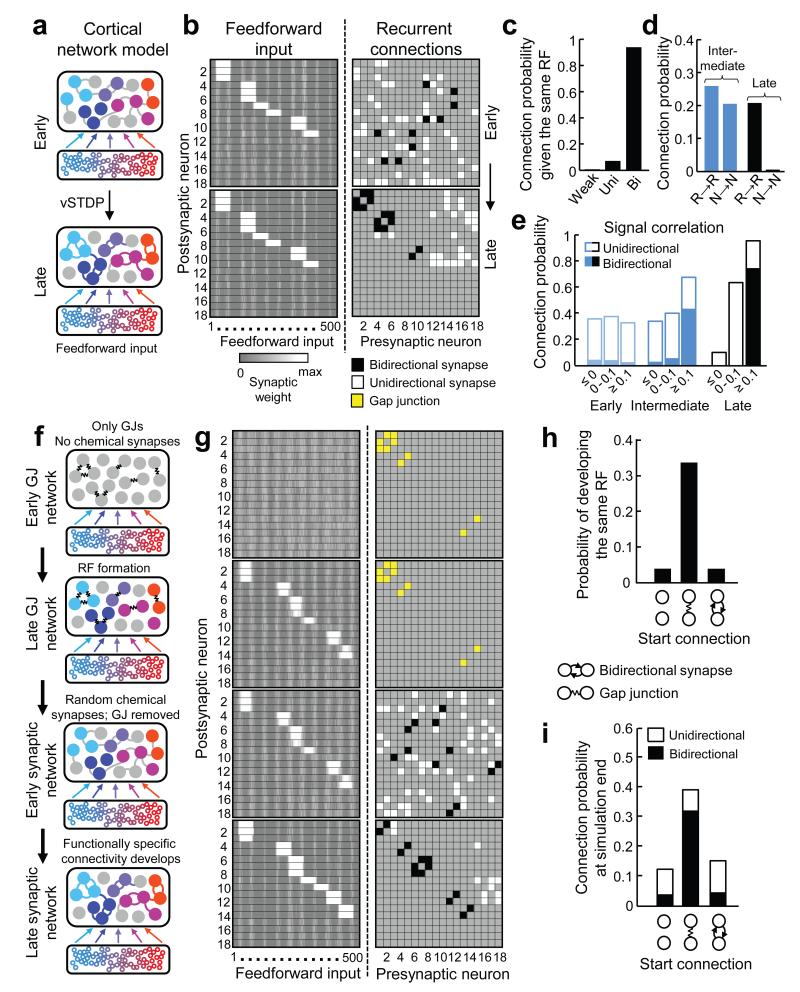
Sensory processing occurs in neocortical microcircuits in which synaptic connectivity is highly structured and excitatory neurons form subnetworks that process related sensory information. However, the developmental mechanisms underlying the formation of functionally organized connectivity in cortical microcircuits remain unknown. Here we directly relate patterns of excitatory synaptic connectivity to visual response properties of neighbouring layer 2/3 pyramidal neurons in mouse visual cortex at different postnatal ages, using two-photon calcium imaging in vivo and multiple whole-cell recordings in vitro. Although neural responses were already highly selective for visual stimuli at eye opening, neurons responding to similar visual features were not yet preferentially connected, indicating that the emergence of feature selectivity does not depend on the precise arrangement of local synaptic connections. After eye opening, local connectivity reorganized extensively: more connections formed selectively between neurons with similar visual responses and connections were eliminated between visually unresponsive neurons, but the overall connectivity rate did not change. We propose a sequential model of cortical microcircuit development based on activity-dependent mechanisms of plasticity whereby neurons first acquire feature preference by selecting feedforward inputs before the onset of sensory experience--a process that may be facilitated by early electrical coupling between neuronal subsets--and then patterned input drives the formation of functional subnetworks through a redistribution of recurrent synaptic connections.
Figures
Comment in
-
Visual processing: eye-opening reorganization.Nat Rev Neurosci. 2013 May;14(5):309. doi: 10.1038/nrn3501. Nat Rev Neurosci. 2013. PMID: 23598725 No abstract available.
References
Methods References
-
- Nimmerjahn A, Kirchhoff F, Kerr JND, Helmchen F. Sulforhodamine 101 as a specific marker of astroglia in the neocortex in vivo. Nat. Methods. 2004;1:31–37. - PubMed
-
- Brainard DH. The Psychophysics Toolbox. Spat Vis. 1997;10:433–436. - PubMed
-
- Pelli DG. The VideoToolbox software for visual psychophysics: transforming numbers into movies. Spat Vis. 1997;10:437–442. - PubMed
