

Cyclic AMP-dependent protein lysine acylation in mycobacteria regulates fatty acid and propionate metabolism
- PMID: 23553634
- PMCID: PMC3656268
- DOI: 10.1074/jbc.M113.463992
Cyclic AMP-dependent protein lysine acylation in mycobacteria regulates fatty acid and propionate metabolism
Erratum in
-
Correction: Cyclic AMP-dependent protein lysine acylation in mycobacteria regulates fatty acid and propionate metabolism.J Biol Chem. 2019 Jul 12;294(28):11046. doi: 10.1074/jbc.AAC119.009859. J Biol Chem. 2019. PMID: 31300591 Free PMC article. No abstract available.
Abstract
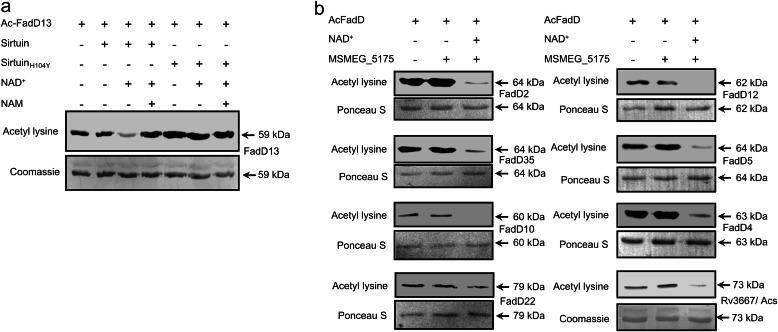
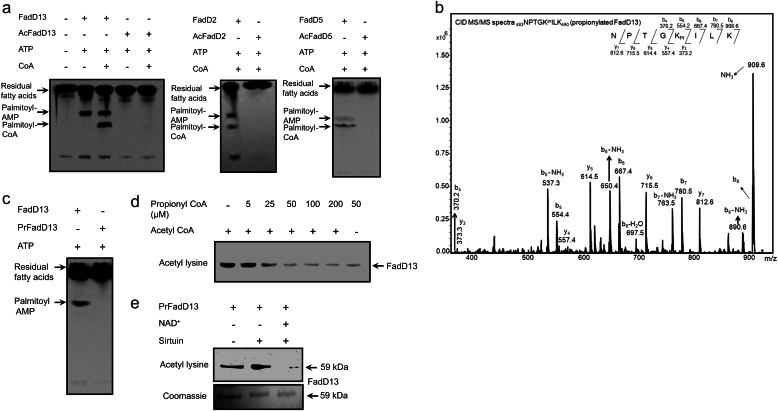
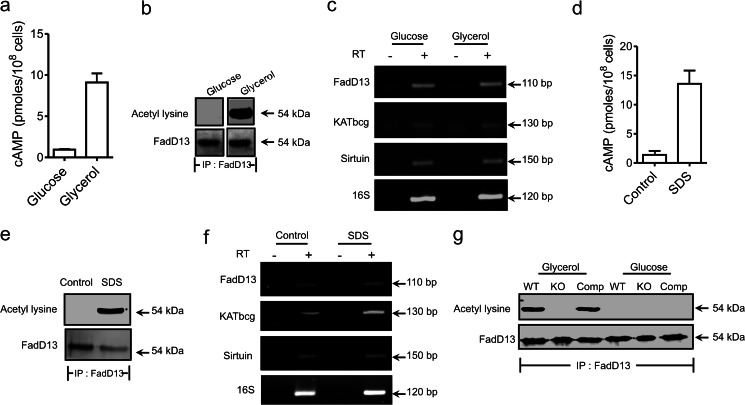
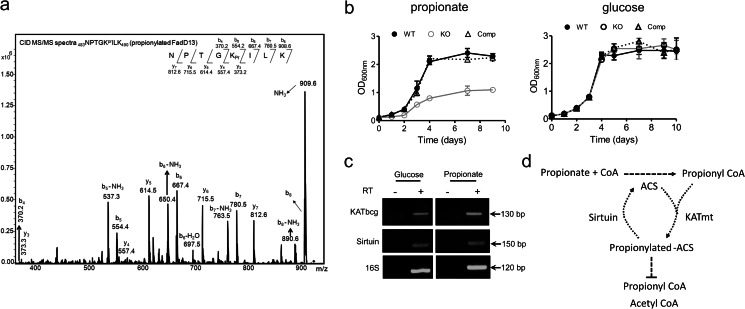
Acetylation of lysine residues is a posttranslational modification that is used by both eukaryotes and prokaryotes to regulate a variety of biological processes. Here we identify multiple substrates for the cAMP-dependent protein lysine acetyltransferase from Mycobacterium tuberculosis (KATmt). We demonstrate that a catalytically important lysine residue in a number of FadD (fatty acyl CoA synthetase) enzymes is acetylated by KATmt in a cAMP-dependent manner and that acetylation inhibits the activity of FadD enzymes. A sirtuin-like enzyme can deacetylate multiple FadDs, thus completing the regulatory cycle. Using a strain deleted for the KATmt ortholog in Mycobacterium bovis Bacillus Calmette-Guérin (BCG), we show for the first time that acetylation is dependent on intracellular cAMP levels. KATmt can utilize propionyl CoA as a substrate and, therefore, plays a critical role in alleviating propionyl CoA toxicity in mycobacteria by inactivating acyl CoA synthetase (ACS). The precision by which mycobacteria can regulate the metabolism of fatty acids in a cAMP-dependent manner appears to be unparalleled in other biological organisms and is ideally suited to adapt to the complex environment that pathogenic mycobacteria experience in the host.
Keywords: Acetyl Coenzyme A; Actinobacteria; Fatty Acid Metabolism; Mass Spectrometry (MS); Mycobacteria; Prokaryotic Signal Transduction; Protein Acylation.
Figures

Similar articles
-
cAMP-regulated protein lysine acetylases in mycobacteria.J Biol Chem. 2010 Aug 6;285(32):24313-23. doi: 10.1074/jbc.M110.118398. Epub 2010 May 27. J Biol Chem. 2010. PMID: 20507997 Free PMC article.
-
Reversible acetylation and inactivation of Mycobacterium tuberculosis acetyl-CoA synthetase is dependent on cAMP.Biochemistry. 2011 Jul 5;50(26):5883-92. doi: 10.1021/bi200156t. Epub 2011 Jun 10. Biochemistry. 2011. PMID: 21627103 Free PMC article.
-
Mycobacterial phenolic glycolipid synthesis is regulated by cAMP-dependent lysine acylation of FadD22.Microbiology (Reading). 2017 Mar;163(3):373-382. doi: 10.1099/mic.0.000440. Epub 2017 Mar 29. Microbiology (Reading). 2017. PMID: 28141495
-
Cyclic AMP Signaling in Mycobacteria.Microbiol Spectr. 2014 Apr;2(2). doi: 10.1128/microbiolspec.MGM2-0011-2013. Microbiol Spectr. 2014. PMID: 26105822 Review.
-
Yeast acyl-CoA synthetases at the crossroads of fatty acid metabolism and regulation.Biochim Biophys Acta. 2007 Mar;1771(3):286-98. doi: 10.1016/j.bbalip.2006.05.003. Epub 2006 May 16. Biochim Biophys Acta. 2007. PMID: 16798075 Review.
Cited by
-
Mechanism and regulation of mycobactin fatty acyl-AMP ligase FadD33.J Biol Chem. 2013 Sep 27;288(39):28116-25. doi: 10.1074/jbc.M113.495549. Epub 2013 Aug 9. J Biol Chem. 2013. PMID: 23935107 Free PMC article.
-
The adenylyl cyclase Rv2212 modifies the proteome and infectivity of Mycobacterium bovis BCG.Folia Microbiol (Praha). 2015 Jan;60(1):21-31. doi: 10.1007/s12223-014-0335-1. Epub 2014 Jul 21. Folia Microbiol (Praha). 2015. PMID: 25038956
-
An acyl-CoA synthetase in Mycobacterium tuberculosis involved in triacylglycerol accumulation during dormancy.PLoS One. 2014 Dec 9;9(12):e114877. doi: 10.1371/journal.pone.0114877. eCollection 2014. PLoS One. 2014. PMID: 25490545 Free PMC article.
-
Post-translational Succinylation of Mycobacterium tuberculosis Enoyl-CoA Hydratase EchA19 Slows Catalytic Hydration of Cholesterol Catabolite 3-Oxo-chol-4,22-diene-24-oyl-CoA.ACS Infect Dis. 2020 Aug 14;6(8):2214-2224. doi: 10.1021/acsinfecdis.0c00329. Epub 2020 Jul 27. ACS Infect Dis. 2020. PMID: 32649175 Free PMC article.
-
Novel proteome and acetylome of Bemisia tabaci Q in response to Cardinium infection.BMC Genomics. 2018 Jul 5;19(1):523. doi: 10.1186/s12864-018-4907-3. BMC Genomics. 2018. PMID: 29976144 Free PMC article.
References
-
- Cole S. T., Brosch R., Parkhill J., Garnier T., Churcher C., Harris D., Gordon S. V., Eiglmeier K., Gas S., Barry C. E., 3rd, Tekaia F., Badcock K., Basham D., Brown D., Chillingworth T., Connor R., Davies R., Devlin K., Feltwell T., Gentles S., Hamlin N., Holroyd S., Hornsby T., Jagels K., Krogh A., McLean J., Moule S., Murphy L., Oliver K., Osborne J., Quail M. A., Rajandream M. A., Rogers J., Rutter S., Seeger K., Skelton J., Squares R., Squares S., Sulston J. E., Taylor K., Whitehead S., Barrell B. G. (1998) Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393, 537–544 - PubMed
-
- McCue L. A., McDonough K. A., Lawrence C. E. (2000) Functional classification of cNMP-binding proteins and nucleotide cyclases with implications for novel regulatory pathways in Mycobacterium tuberculosis. Genome Res. 10, 204–219 - PubMed
-
- Shenoy A. R., Visweswariah S. S. (2006) New messages from old messengers. cAMP and mycobacteria. Trends Microbiol. 14, 543–550 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
