

HIV-1 Tat C modulates expression of miRNA-101 to suppress VE-cadherin in human brain microvascular endothelial cells
- PMID: 23554480
- PMCID: PMC6618916
- DOI: 10.1523/JNEUROSCI.4796-12.2013
HIV-1 Tat C modulates expression of miRNA-101 to suppress VE-cadherin in human brain microvascular endothelial cells
Abstract
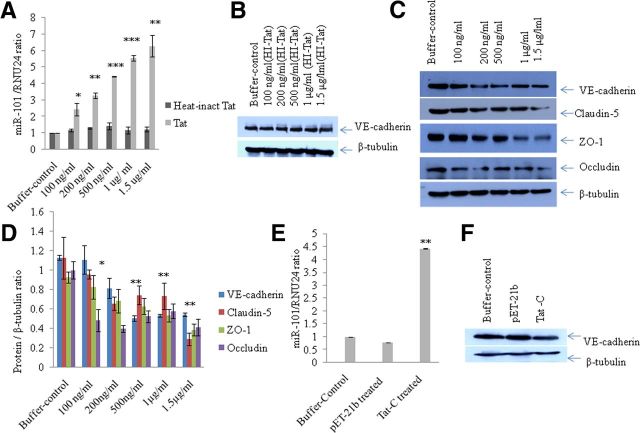
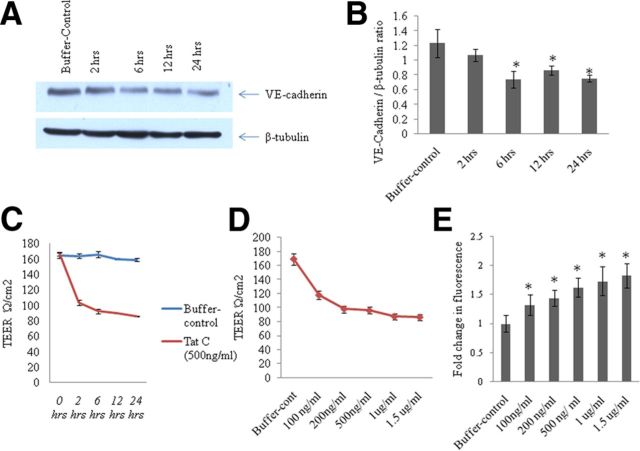
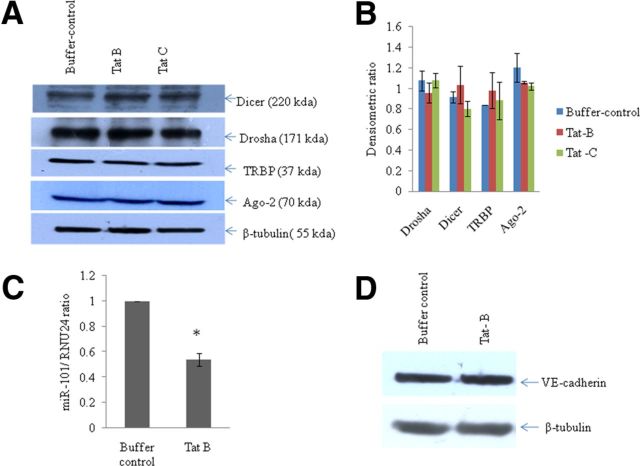
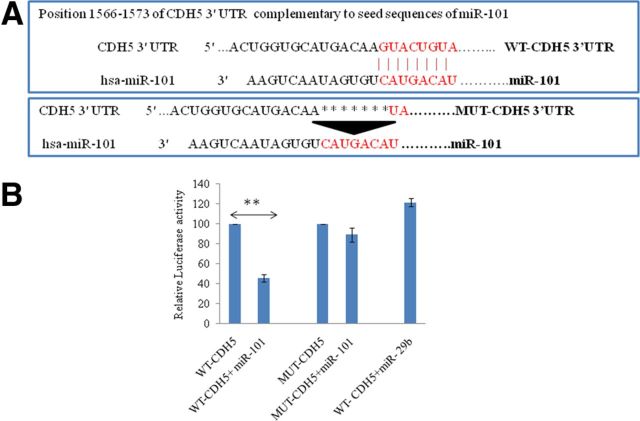
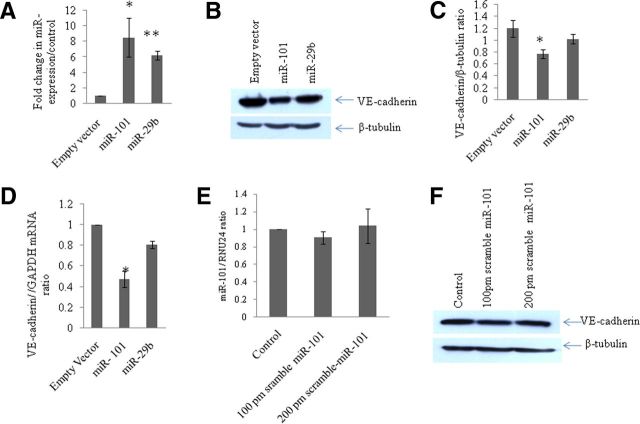
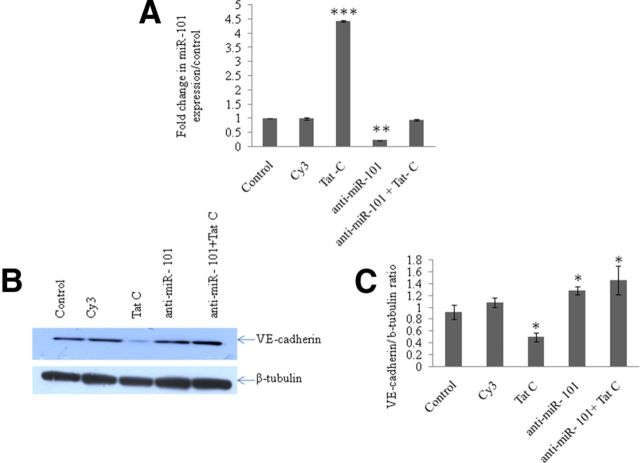
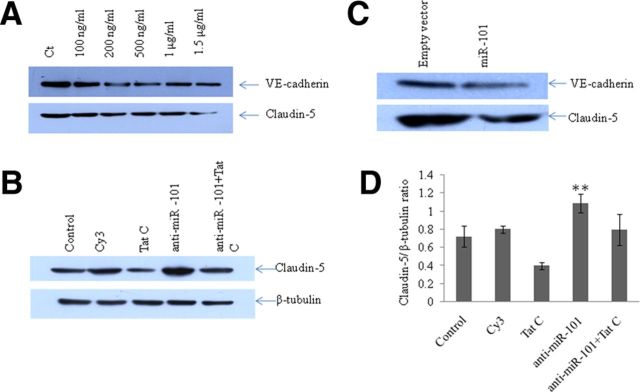
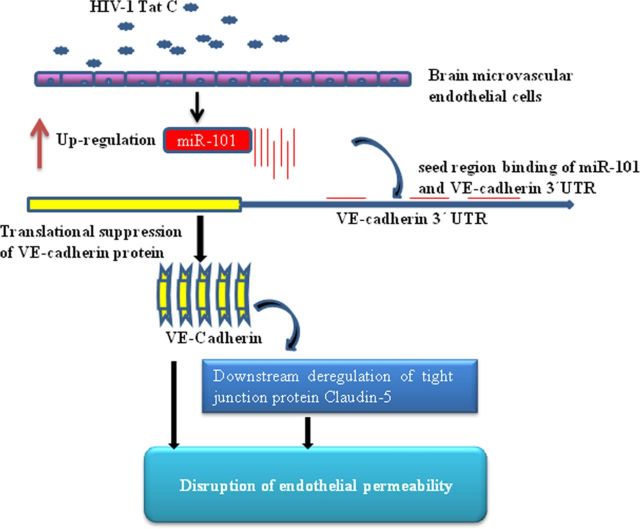
HIV-1 infection leads to the development of HIV-associated neurological disorders. The HIV-1 Tat protein has been reported to exert an adverse effect on blood-brain barrier integrity and permeability. Perturbation in permeability is mainly caused by disruptions in adherens junctions and tight junction proteins. We have identified HIV-1 Tat C-induced disruption of VE-cadherin mediated by miRNA-101 in human brain microvascular endothelial cells (BMVECs). HIV-1 Tat C increased the expression of miR-101, which led to downregulation of VE-cadherin. Overexpression of miR-101 resulted into the suppression of VE-cadherin. Inhibition of miR-101 by the miRNA inhibitor enhanced the expression of VE-cadherin. We have demonstrated that VE-cadherin is a direct target of miR-101 using a luciferase reporter assay, which showed that mutated VE-cadherin 3'UTR and miR-101 cotransfection did not change luciferase activity. By overexpression and knockdown of miR-101, we have demonstrated that the expression level of claudin-5 is governed by the expression of VE-cadherin. These findings demonstrate a novel mechanism for the regulation of barrier permeability by miR-101 via posttranscriptional regulation of VE-cadherin in human BMVECs exposed to the HIV-1 Tat C protein.
Figures
Similar articles
-
HIV-1 Tat C phosphorylates VE-cadherin complex and increases human brain microvascular endothelial cell permeability.BMC Neurosci. 2014 Jun 26;15:80. doi: 10.1186/1471-2202-15-80. BMC Neurosci. 2014. PMID: 24965120 Free PMC article.
-
Zika Virus NS1 Suppresses VE-Cadherin and Claudin-5 via hsa-miR-101-3p in Human Brain Microvascular Endothelial Cells.Mol Neurobiol. 2021 Dec;58(12):6290-6303. doi: 10.1007/s12035-021-02548-x. Epub 2021 Sep 6. Mol Neurobiol. 2021. PMID: 34487317
-
MicroRNA Regulation of Endothelial Junction Proteins and Clinical Consequence.Mediators Inflamm. 2016;2016:5078627. doi: 10.1155/2016/5078627. Epub 2016 Nov 24. Mediators Inflamm. 2016. PMID: 27999452 Free PMC article. Review.
-
Hypoxia-Induced miR-101 Impairs Endothelial Barrier Integrity Through Altering VE-Cadherin and Claudin-5.Mol Neurobiol. 2024 Mar;61(3):1807-1817. doi: 10.1007/s12035-023-03662-8. Epub 2023 Sep 30. Mol Neurobiol. 2024. PMID: 37776496
-
Dynamic Regulation of Vascular Permeability by Vascular Endothelial Cadherin-Mediated Endothelial Cell-Cell Junctions.J Nippon Med Sch. 2017;84(4):148-159. doi: 10.1272/jnms.84.148. J Nippon Med Sch. 2017. PMID: 28978894 Review.
Cited by
-
The characterization of extracellular vesicles-derived microRNAs in Thai malaria patients.Malar J. 2020 Aug 10;19(1):285. doi: 10.1186/s12936-020-03360-z. Malar J. 2020. PMID: 32778117 Free PMC article.
-
miRNAs Participate in MS Pathological Processes and Its Therapeutic Response.Mediators Inflamm. 2016;2016:4578230. doi: 10.1155/2016/4578230. Epub 2016 Mar 17. Mediators Inflamm. 2016. PMID: 27073296 Free PMC article. Review.
-
Non-coding RNAs in cerebral endothelial pathophysiology: emerging roles in stroke.Neurochem Int. 2014 Nov;77:9-16. doi: 10.1016/j.neuint.2014.03.013. Epub 2014 Apr 3. Neurochem Int. 2014. PMID: 24704794 Free PMC article. Review.
-
HIV-1 Tat C phosphorylates VE-cadherin complex and increases human brain microvascular endothelial cell permeability.BMC Neurosci. 2014 Jun 26;15:80. doi: 10.1186/1471-2202-15-80. BMC Neurosci. 2014. PMID: 24965120 Free PMC article.
-
Tight junctions and their regulation by non-coding RNAs.Int J Biol Sci. 2021 Jan 31;17(3):712-727. doi: 10.7150/ijbs.45885. eCollection 2021. Int J Biol Sci. 2021. PMID: 33767583 Free PMC article. Review.
References
-
- Alexander JS, Alexander BC, Eppihimer LA, Goodyear N, Haque R, Davis CP, Kalogeris TJ, Carden DL, Zhu YN, Kevil CG. Inflammatory mediators induce sequestration of VE-cadherin in cultured human endothelial cells. Inflammation. 2000;24:99–113. - PubMed
-
- András IE, Pu H, Tian J, Deli MA, Nath A, Hennig B, Toborek M. Signaling mechanisms of HIV-1 Tat-induced alterations of claudin-5 expression in brain endothelial cells. J Cereb Blood Flow Metab. 2005;25:1159–1170. - PubMed
-
- Andriopoulou P, Navarro P, Zanetti A, Lampugnani MG, Dejana E. Histamine induces tyrosine phosphorylation of endothelial cell-to-cell adherens junctions. Arterioscler Thromb Vasc Biol. 1999;19:2286–2297. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases