

Sucrose ingestion induces rapid AMPA receptor trafficking
- PMID: 23554493
- PMCID: PMC3767387
- DOI: 10.1523/JNEUROSCI.4806-12.2013
Sucrose ingestion induces rapid AMPA receptor trafficking
Abstract
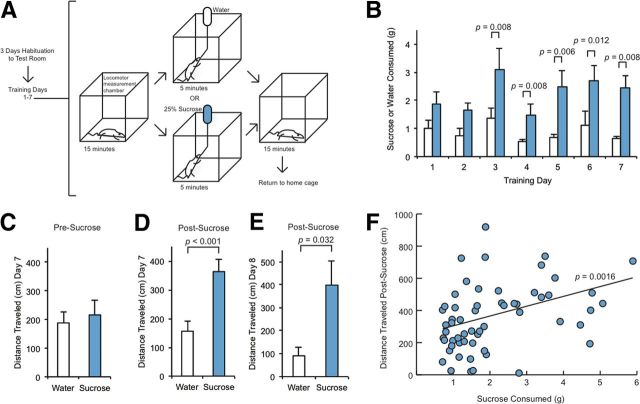
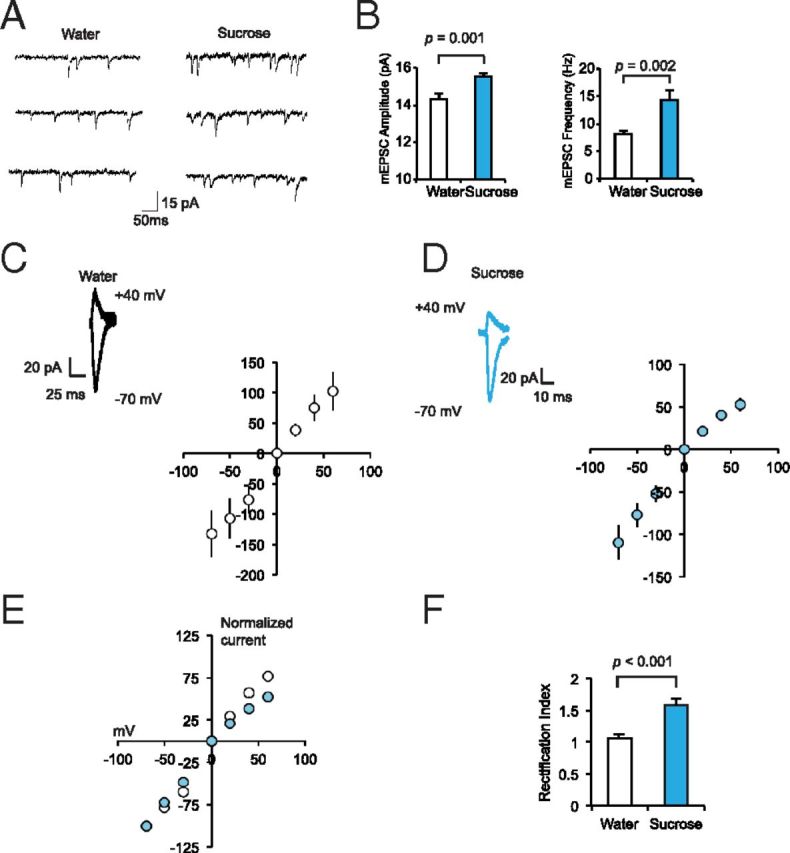
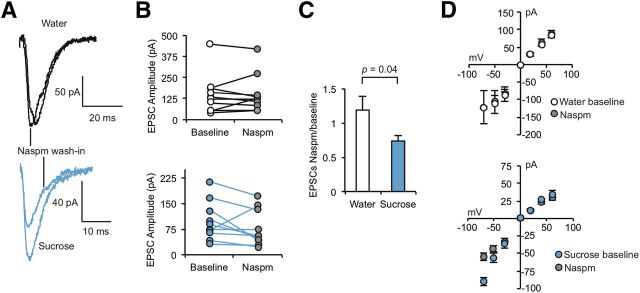
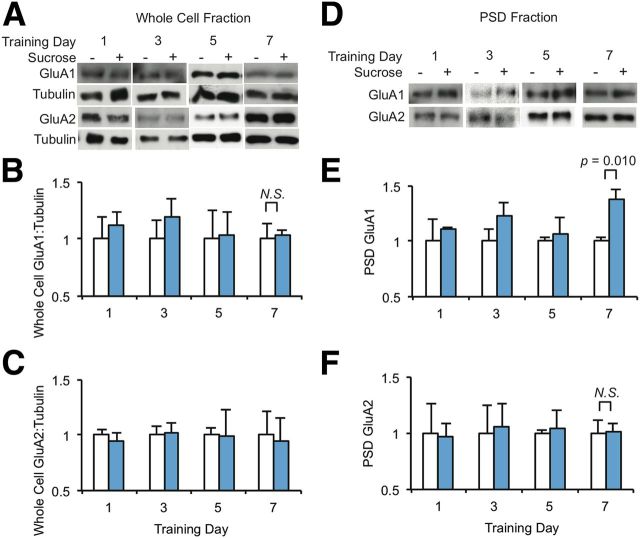
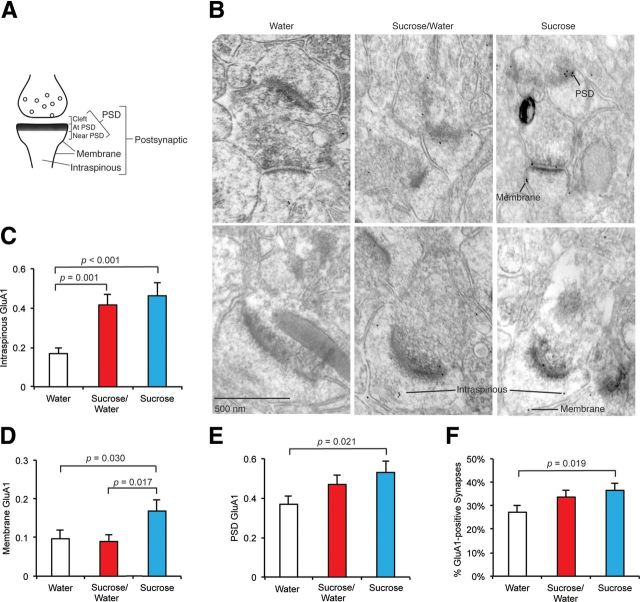
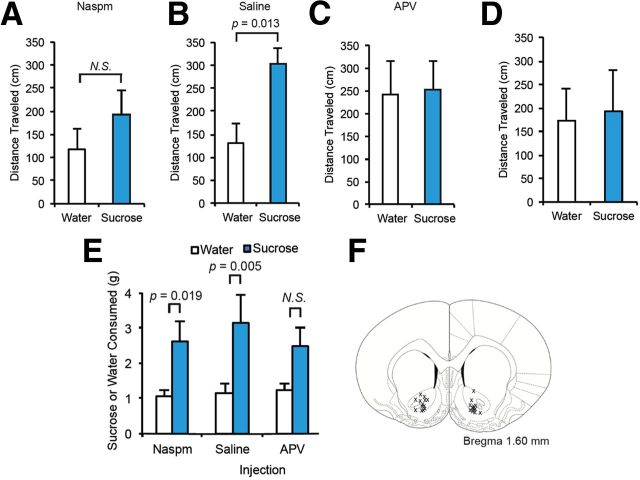
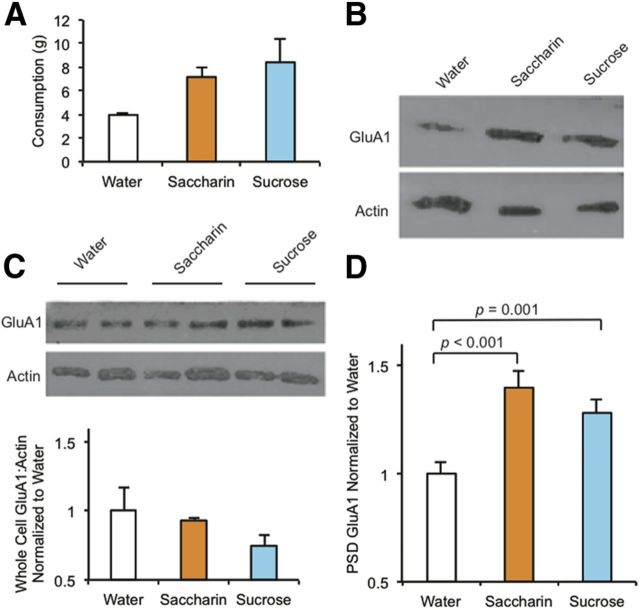
The mechanisms by which natural rewards such as sugar affect synaptic transmission and behavior are largely unexplored. Here, we investigate regulation of nucleus accumbens synapses by sucrose intake. Previous studies have shown that AMPA receptor (AMPAR) trafficking is a major mechanism for regulating synaptic strength, and that in vitro, trafficking of AMPARs containing the GluA1 subunit takes place by a two-step mechanism involving extrasynaptic and then synaptic receptor transport. We report that in rat, repeated daily ingestion of a 25% sucrose solution transiently elevated spontaneous locomotion and potentiated accumbens core synapses through incorporation of Ca(2+)-permeable AMPA receptors (CPARs), which are GluA1-containing, GluA2-lacking AMPARs. Electrophysiological, biochemical, and quantitative electron microscopy studies revealed that sucrose training (7 d) induced a stable (>24 h) intraspinous GluA1 population, and that in these rats a single sucrose stimulus rapidly (5 min) but transiently (<24 h) elevated GluA1 at extrasynaptic sites. CPARs and dopamine D1 receptors were required in vivo for elevated locomotion after sucrose ingestion. Significantly, a 7 d protocol of daily ingestion of a 3% solution of saccharin, a noncaloric sweetener, induced synaptic GluA1 similarly to 25% sucrose ingestion. These findings identify multistep GluA1 trafficking, previously described in vitro, as a mechanism for acute regulation of synaptic transmission in vivo by a natural orosensory reward. Trafficking is stimulated by a chemosensory pathway that is not dependent on the caloric value of sucrose.
Figures
Similar articles
-
Episodic sucrose intake during food restriction increases synaptic abundance of AMPA receptors in nucleus accumbens and augments intake of sucrose following restoration of ad libitum feeding.Neuroscience. 2015 Jun 4;295:58-71. doi: 10.1016/j.neuroscience.2015.03.025. Epub 2015 Mar 20. Neuroscience. 2015. PMID: 25800309 Free PMC article.
-
Food restriction induces synaptic incorporation of calcium-permeable AMPA receptors in nucleus accumbens.Eur J Neurosci. 2017 Mar;45(6):826-836. doi: 10.1111/ejn.13528. Epub 2017 Feb 15. Eur J Neurosci. 2017. PMID: 28112453 Free PMC article.
-
Calcineurin regulates synaptic Ca2+-permeable AMPA receptors in hypothalamic presympathetic neurons via α2δ-1-mediated GluA1/GluA2 assembly.J Physiol. 2024 May;602(10):2179-2197. doi: 10.1113/JP286081. Epub 2024 Apr 17. J Physiol. 2024. PMID: 38630836 Free PMC article.
-
Ca2+-permeable AMPA (α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) receptors and dopamine D1 receptors regulate GluA1 trafficking in striatal neurons.J Biol Chem. 2013 Dec 6;288(49):35297-306. doi: 10.1074/jbc.M113.516690. Epub 2013 Oct 16. J Biol Chem. 2013. PMID: 24133208 Free PMC article.
-
Subunit-specific trafficking mechanisms regulating the synaptic expression of Ca(2+)-permeable AMPA receptors.Semin Cell Dev Biol. 2014 Mar;27:14-22. doi: 10.1016/j.semcdb.2013.12.002. Epub 2013 Dec 15. Semin Cell Dev Biol. 2014. PMID: 24342448 Review.
Cited by
-
Incorporation of inwardly rectifying AMPA receptors at silent synapses during hippocampal long-term potentiation.Philos Trans R Soc Lond B Biol Sci. 2013 Dec 2;369(1633):20130156. doi: 10.1098/rstb.2013.0156. Print 2014 Jan 5. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 24298157 Free PMC article.
-
Linking Cholinergic Interneurons, Synaptic Plasticity, and Behavior during the Extinction of a Cocaine-Context Association.Neuron. 2016 Jun 1;90(5):1071-1085. doi: 10.1016/j.neuron.2016.05.001. Epub 2016 May 19. Neuron. 2016. PMID: 27210555 Free PMC article.
-
Effects of junk-food on food-motivated behavior and nucleus accumbens glutamate plasticity; insights into the mechanism of calcium-permeable AMPA receptor recruitment.Neuropharmacology. 2024 Jan 1;242:109772. doi: 10.1016/j.neuropharm.2023.109772. Epub 2023 Oct 26. Neuropharmacology. 2024. PMID: 37898332 Free PMC article.
-
Homeostatic regulation of reward via synaptic insertion of calcium-permeable AMPA receptors in nucleus accumbens.Physiol Behav. 2020 May 15;219:112850. doi: 10.1016/j.physbeh.2020.112850. Epub 2020 Feb 21. Physiol Behav. 2020. PMID: 32092445 Free PMC article. Review.
-
Episodic sucrose intake during food restriction increases synaptic abundance of AMPA receptors in nucleus accumbens and augments intake of sucrose following restoration of ad libitum feeding.Neuroscience. 2015 Jun 4;295:58-71. doi: 10.1016/j.neuroscience.2015.03.025. Epub 2015 Mar 20. Neuroscience. 2015. PMID: 25800309 Free PMC article.
References
-
- Basar K, Sesia T, Groenewegen H, Steinbusch HW, Visser-Vandewalle V, Temel Y. Nucleus accumbens and impulsivity. Prog Neurobiol. 2010;92:533–557. - PubMed
-
- Boehm J, Kang MG, Johnson RC, Esteban J, Huganir RL, Malinow R. Synaptic incorporation of AMPA receptors during LTP is controlled by a PKC phosphorylation site on GluR1. Neuron. 2006;51:213–225. - PubMed
-
- Boudreau AC, Reimers JM, Milovanovic M, Wolf ME. Cell surface AMPA receptors in the rat nucleus accumbens increase during cocaine withdrawal but internalize after cocaine challenge in association with altered activation of mitogen-activated protein kinases. J Neurosci. 2007;27:10621–10635. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 MH096331/MH/NIMH NIH HHS/United States
- R00 DC009635/DC/NIDCD NIH HHS/United States
- 1R21MH091445-01/MH/NIMH NIH HHS/United States
- F31MH76617-01/MH/NIMH NIH HHS/United States
- P30 EY013079/EY/NEI NIH HHS/United States
- R21 MH091445/MH/NIMH NIH HHS/United States
- P30EY13079/EY/NEI NIH HHS/United States
- R01 DA003956/DA/NIDA NIH HHS/United States
- R01NS061920/NS/NINDS NIH HHS/United States
- T32 DC000063/DC/NIDCD NIH HHS/United States
- 5T32DC000063/DC/NIDCD NIH HHS/United States
- DA003956/DA/NIDA NIH HHS/United States
- F31 MH076617/MH/NIMH NIH HHS/United States
- R01 NS061920/NS/NINDS NIH HHS/United States
- T32 MH019524/MH/NIMH NIH HHS/United States
- K99 DC009635/DC/NIDCD NIH HHS/United States
- R56 DA003956/DA/NIDA NIH HHS/United States
- DC009635/DC/NIDCD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous