

Dissociation between dorsal and ventral hippocampal theta oscillations during decision-making
- PMID: 23554502
- PMCID: PMC6618918
- DOI: 10.1523/JNEUROSCI.2915-12.2013
Dissociation between dorsal and ventral hippocampal theta oscillations during decision-making
Abstract
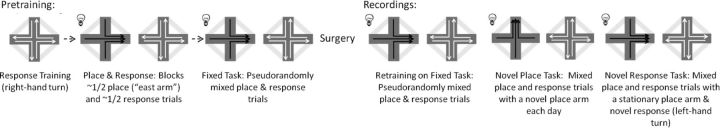
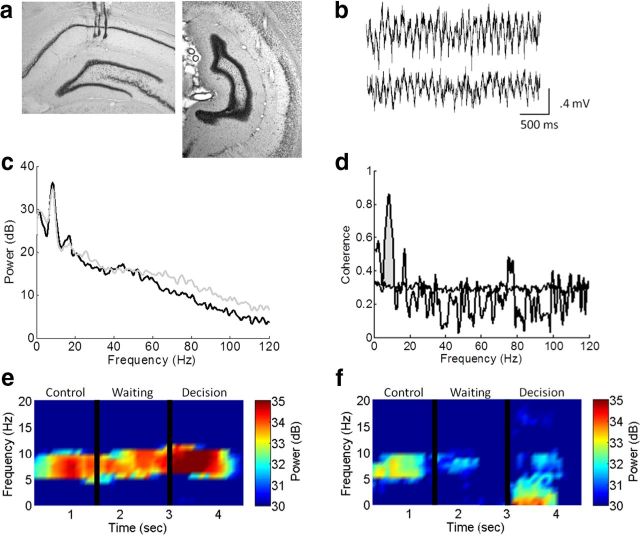
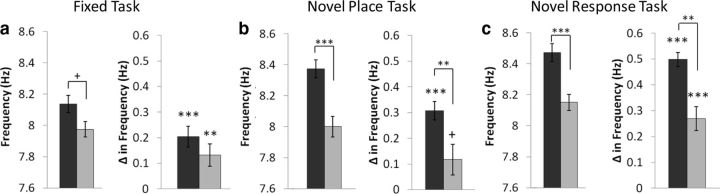
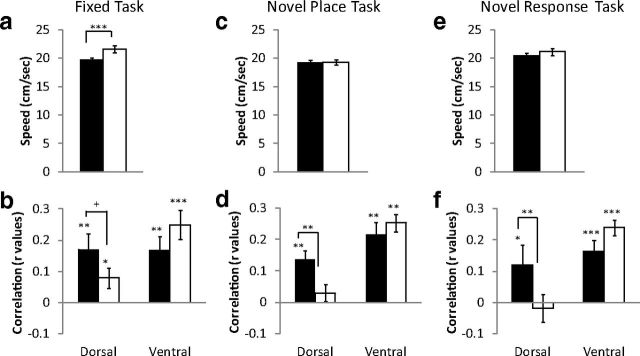
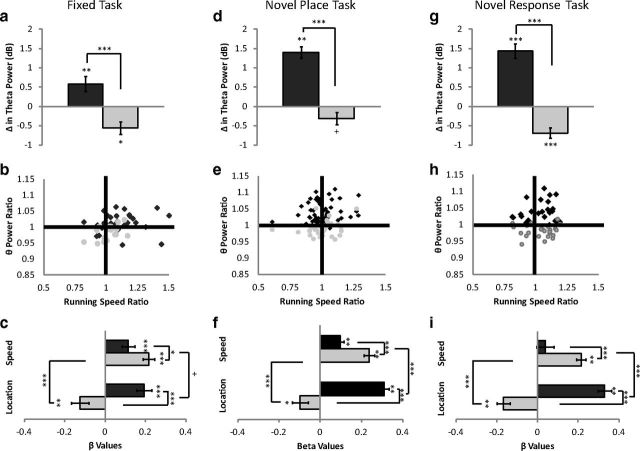
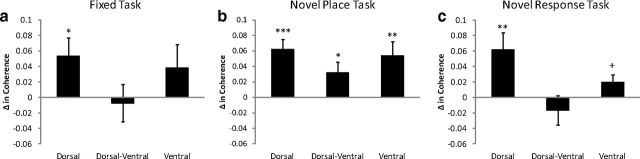
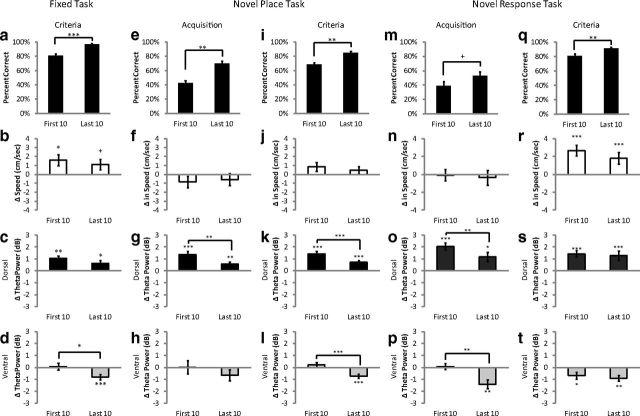
Hippocampal theta oscillations are postulated to support mnemonic processes in humans and rodents. Theta oscillations facilitate encoding and spatial navigation, but to date, it has been difficult to dissociate the effects of volitional movement from the cognitive demands of a task. Therefore, we examined whether volitional movement or cognitive demands exerted a greater modulating factor over theta oscillations during decision-making. Given the anatomical, electrophysiological, and functional dissociations along the dorsal-ventral axis, theta oscillations were simultaneously recorded in the dorsal and ventral hippocampus in rats trained to switch between place and motor-response strategies. Stark differences in theta characteristics were found between the dorsal and ventral hippocampus in frequency, power, and coherence. Theta power increased in the dorsal, but decreased in the ventral hippocampus, during the decision-making epoch. Interestingly, the relationship between running speed and theta power was uncoupled during the decision-making epoch, a phenomenon limited to the dorsal hippocampus. Theta frequency increased in both the dorsal and ventral hippocampus during the decision epoch, although this effect was greater in the dorsal hippocampus. Despite these differences, ventral hippocampal theta was responsive to the navigation task; theta frequency, power, and coherence were all affected by cognitive demands. Theta coherence increased within the dorsal hippocampus during the decision-making epoch on all three tasks. However, coherence selectively increased throughout the hippocampus (dorsal to ventral) on the task with new hippocampal learning. Interestingly, most results were consistent across tasks, regardless of hippocampal-dependent learning. These data indicate increased integration and cooperation throughout the hippocampus during information processing.
Figures
References
-
- Aigner TG, Mishkin M. The effects of physostigmine and scopolamine on recognition memory in monkeys. Behav Neural Biol. 1986;45:81–87. - PubMed
-
- Amaral D, Lavenex P. The hippocampus book. Oxford: Oxford UP; 2007. Hippocampal neuroanatomy.
-
- Benchenane K, Peyrache A, Khamassi M, Tierney PL, Gioanni Y, Battaglia FP, Wiener SI. Coherent theta oscillations and reorganization of spike timing in the hippocampal-prefrontal network upon leaning. Neuron. 2010;66:921–936. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources