

Functional characterization of the extraclassical receptive field in macaque V1: contrast, orientation, and temporal dynamics
- PMID: 23554504
- PMCID: PMC3675885
- DOI: 10.1523/JNEUROSCI.4155-12.2013
Functional characterization of the extraclassical receptive field in macaque V1: contrast, orientation, and temporal dynamics
Abstract
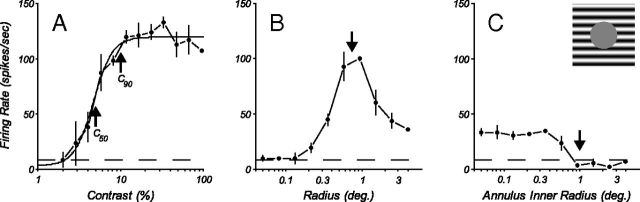
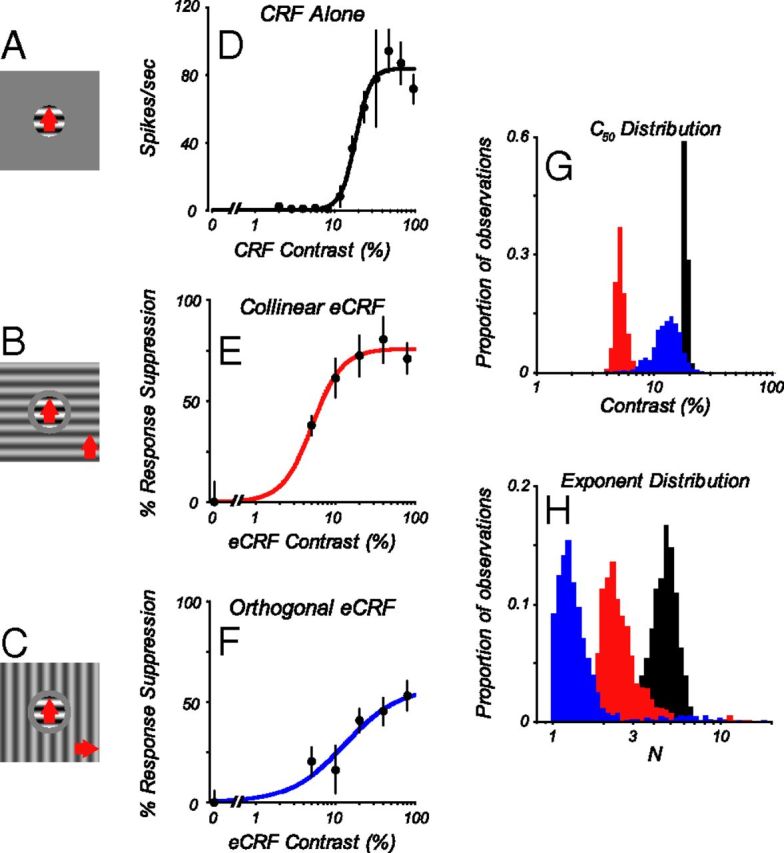
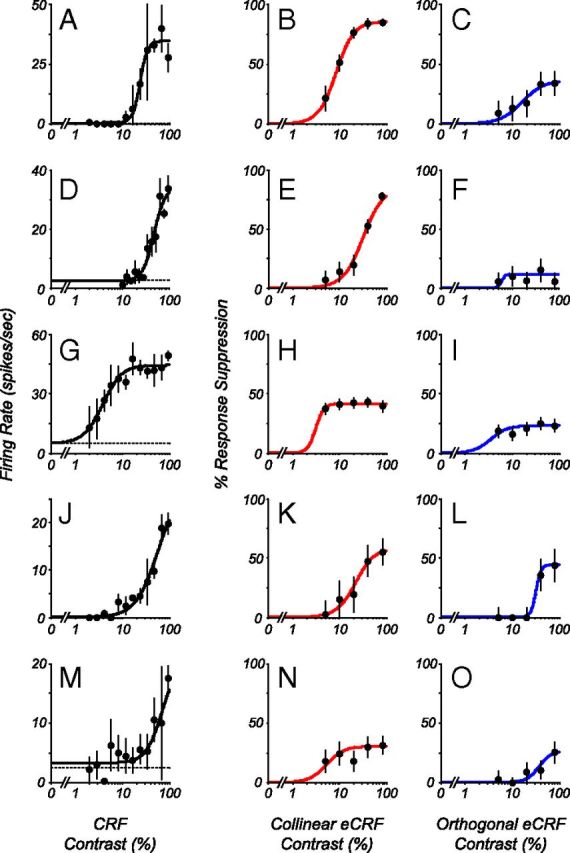
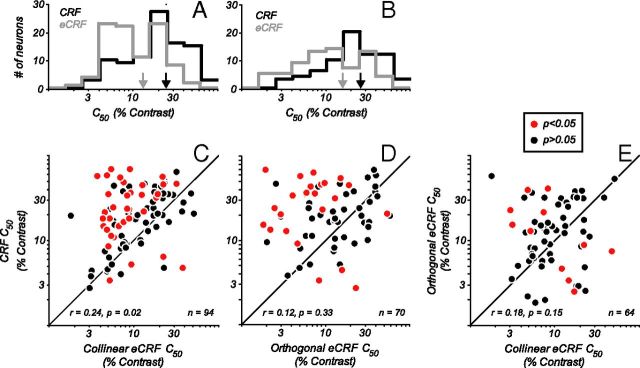
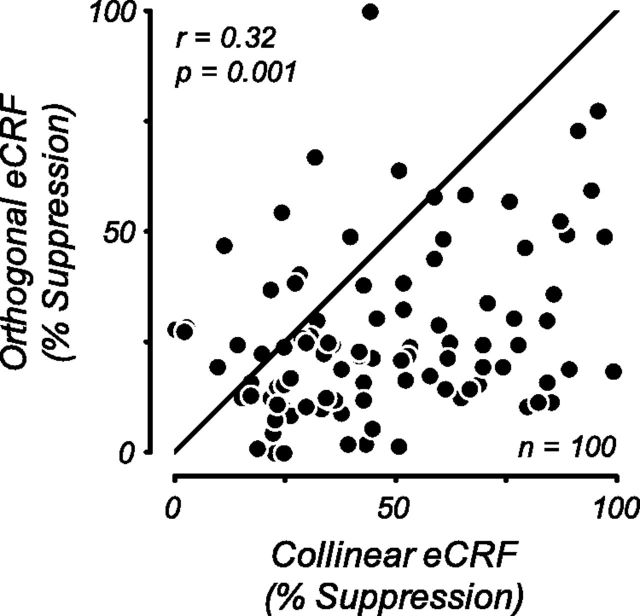
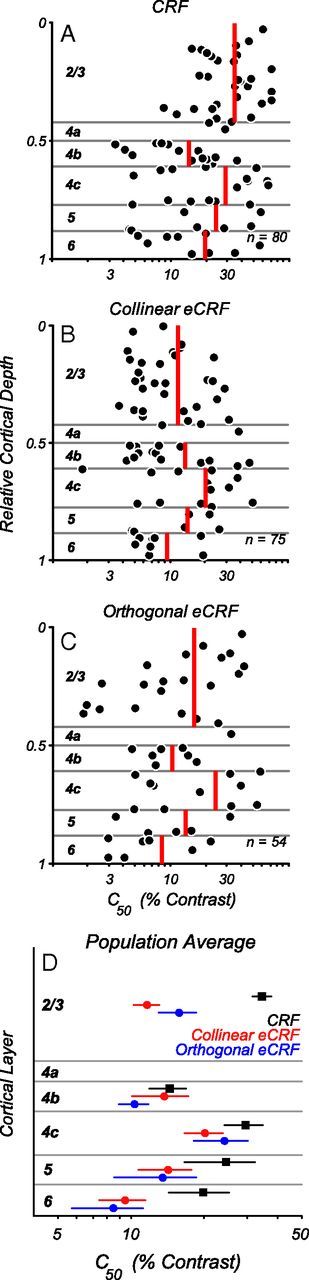
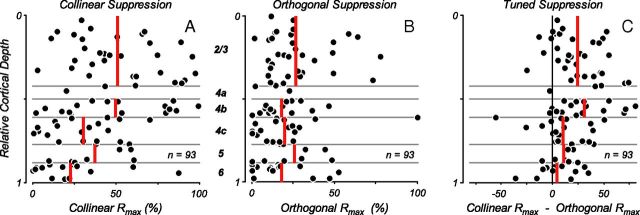
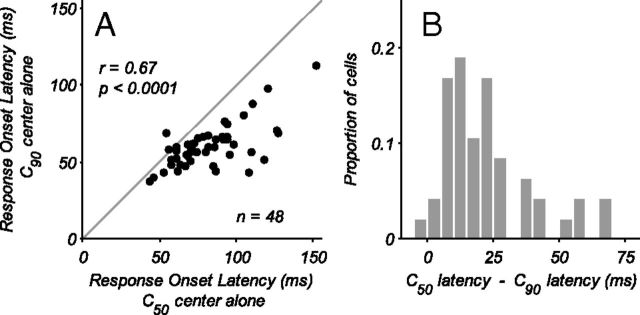
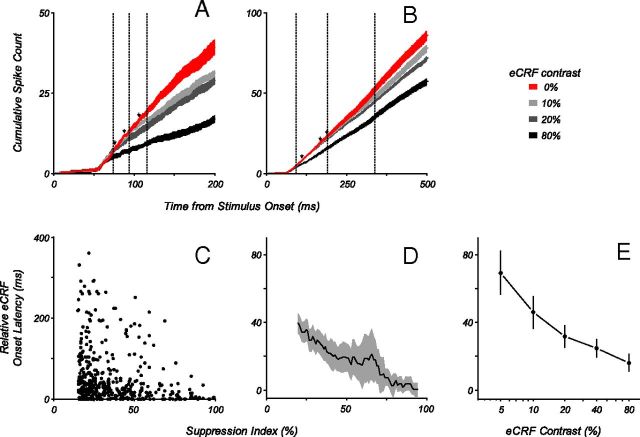
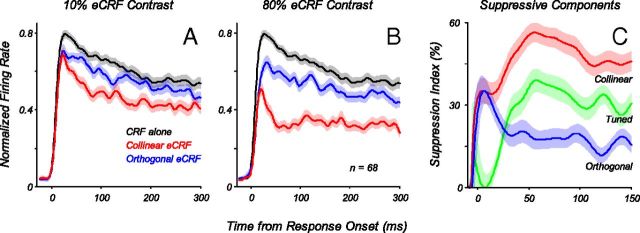
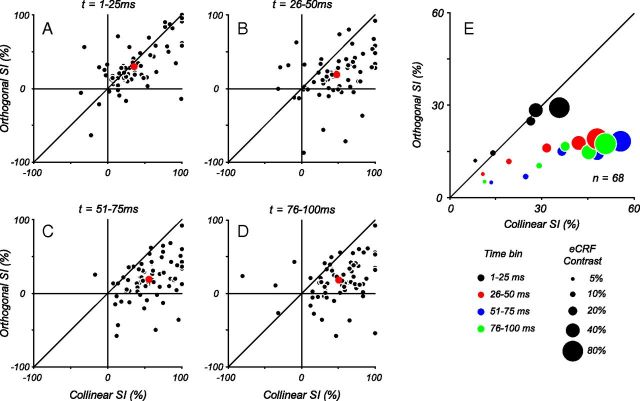
Neurons in primary visual cortex, V1, very often have extraclassical receptive fields (eCRFs). The eCRF is defined as the region of visual space where stimuli cannot elicit a spiking response but can modulate the response of a stimulus in the classical receptive field (CRF). We investigated the dependence of the eCRF on stimulus contrast and orientation in macaque V1 cells for which the laminar location was determined. The eCRF was more sensitive to contrast than the CRF across the whole population of V1 cells with the greatest contrast differential in layer 2/3. We confirmed that many V1 cells experience stronger suppression for collinear than orthogonal stimuli in the eCRF. Laminar analysis revealed that the predominant bias for collinear suppression was found in layers 2/3 and 4b. The laminar pattern of contrast and orientation dependence suggests that eCRF suppression may derive from different neural circuits in different layers, and may be comprised of two distinct components: orientation-tuned and untuned suppression. On average tuned suppression was delayed by ∼25 ms compared with the onset of untuned suppression. Therefore, response modulation by the eCRF develops dynamically and rapidly in time.
Figures
Similar articles
-
Distinct spatiotemporal mechanisms underlie extra-classical receptive field modulation in macaque V1 microcircuits.Elife. 2020 May 27;9:e54264. doi: 10.7554/eLife.54264. Elife. 2020. PMID: 32458798 Free PMC article.
-
Orientation tuning of surround suppression in lateral geniculate nucleus and primary visual cortex of cat.Neuroscience. 2007 Nov 23;149(4):962-75. doi: 10.1016/j.neuroscience.2007.08.001. Epub 2007 Aug 9. Neuroscience. 2007. PMID: 17945429
-
Surround suppression sharpens orientation tuning in the cat primary visual cortex.Eur J Neurosci. 2009 Mar;29(5):1035-46. doi: 10.1111/j.1460-9568.2009.06645.x. Eur J Neurosci. 2009. PMID: 19291228
-
Surround suppression supports second-order feature encoding by macaque V1 and V2 neurons.Vision Res. 2014 Nov;104:24-35. doi: 10.1016/j.visres.2014.10.004. Epub 2014 Oct 23. Vision Res. 2014. PMID: 25449336 Free PMC article. Review.
-
The contribution of vertical and horizontal connections to the receptive field center and surround in V1.Neural Netw. 2004 Jun-Jul;17(5-6):681-93. doi: 10.1016/j.neunet.2004.05.002. Neural Netw. 2004. PMID: 15288892 Review.
Cited by
-
LFP polarity changes across cortical and eccentricity in primary visual cortex.Front Neurosci. 2023 Feb 27;17:1138602. doi: 10.3389/fnins.2023.1138602. eCollection 2023. Front Neurosci. 2023. PMID: 36922925 Free PMC article.
-
Selective tuning for contrast in macaque area V4.J Neurosci. 2013 Nov 20;33(47):18583-96. doi: 10.1523/JNEUROSCI.3465-13.2013. J Neurosci. 2013. PMID: 24259580 Free PMC article.
-
Spatial context non-uniformly modulates inter-laminar information flow in the primary visual cortex.Neuron. 2024 Dec 18;112(24):4081-4095.e5. doi: 10.1016/j.neuron.2024.09.021. Epub 2024 Oct 22. Neuron. 2024. PMID: 39442514
-
The effects of orientation and attention during surround suppression of small image features: A 7 Tesla fMRI study.J Vis. 2016 Aug 1;16(10):19. doi: 10.1167/16.10.19. J Vis. 2016. PMID: 27565016 Free PMC article.
-
Dynamic Contextual Modulation in Superior Colliculus of Awake Mouse.eNeuro. 2020 Sep 15;7(5):ENEURO.0131-20.2020. doi: 10.1523/ENEURO.0131-20.2020. Print 2020 Sep/Oct. eNeuro. 2020. PMID: 32868308 Free PMC article.
References
-
- Allman J, Miezin F, McGuinness E. Stimulus specific responses from beyond the classical receptive field: neurophysiological mechanisms for local-global comparisons in visual neurons. Annu Rev Neurosci. 1985;8:407–430. - PubMed
-
- Ayaz A, Chance FS. Gain modulation of neuronal responses by subtractive and divisive mechanisms of inhibition. J Neurophysiol. 2009;101:958–968. - PubMed
-
- Baker DH, Graf EW. Contextual effects in speed perception may occur at an early stage of processing. Vision Res. 2010;50:193–201. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources