

Positively charged residues are the major determinants of ribosomal velocity
- PMID: 23554576
- PMCID: PMC3595205
- DOI: 10.1371/journal.pbio.1001508
Positively charged residues are the major determinants of ribosomal velocity
Abstract
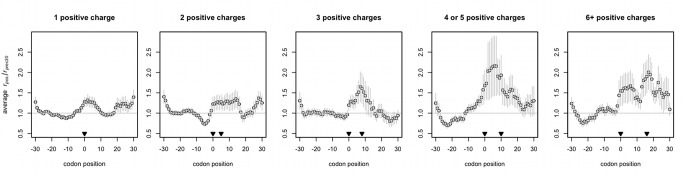
Both for understanding mechanisms of disease and for the design of transgenes, it is important to understand the determinants of ribosome velocity, as changes in the rate of translation are important for protein folding, error attenuation, and localization. While there is great variation in ribosomal occupancy along even a single transcript, what determines a ribosome's occupancy is unclear. We examine this issue using data from a ribosomal footprinting assay in yeast. While codon usage is classically considered a major determinant, we find no evidence for this. By contrast, we find that positively charged amino acids greatly retard ribosomes downstream from where they are encoded, consistent with the suggestion that positively charged residues interact with the negatively charged ribosomal exit tunnel. Such slowing is independent of and greater than the average effect owing to mRNA folding. The effect of charged amino acids is additive, with ribosomal occupancy well-predicted by a linear fit to the density of positively charged residues. We thus expect that a translated poly-A tail, encoding for positively charged lysines regardless of the reading frame, would act as a sandtrap for the ribosome, consistent with experimental data.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
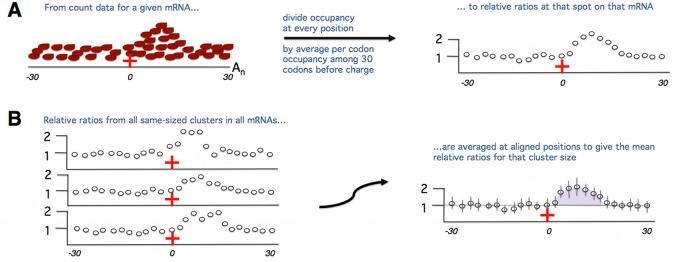
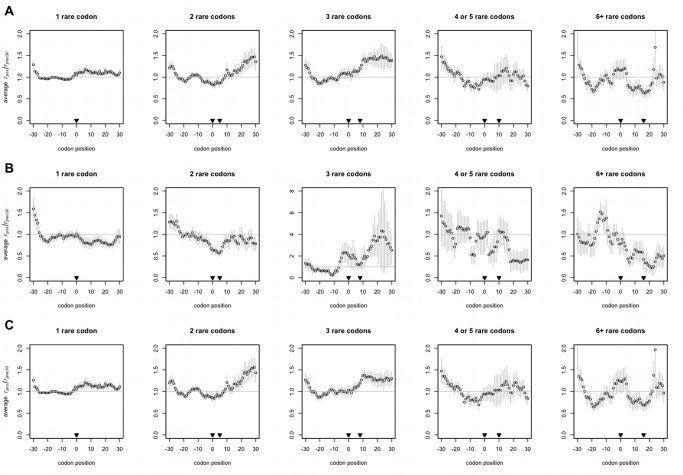
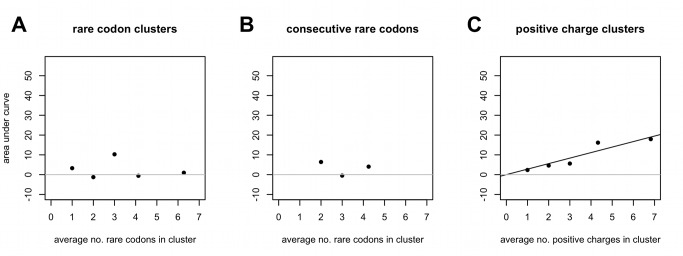
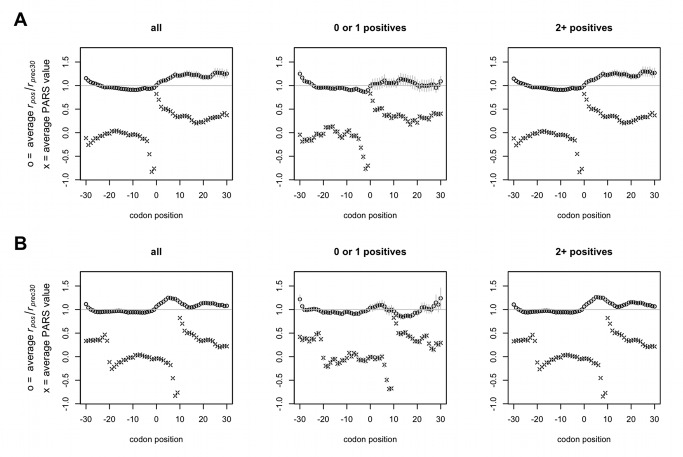
Comment in
-
Gene regulation: In charge of translation.Nat Rev Genet. 2013 May;14(5):304. doi: 10.1038/nrg3478. Epub 2013 Apr 4. Nat Rev Genet. 2013. PMID: 23552216 No abstract available.
-
Positive charges put the brakes on ribosomes.PLoS Biol. 2013;11(3):e1001509. doi: 10.1371/journal.pbio.1001509. Epub 2013 Mar 12. PLoS Biol. 2013. PMID: 23554577 Free PMC article. No abstract available.
References
-
- Randall LL, Josefsson LG, Hardy SJ (1980) Novel intermediates in the synthesis of maltose-binding protein in Escherichia coli. Eur J Biochem 107: 375–379. - PubMed
-
- Siller E, DeZwaan DC, Anderson JF, Freeman BC, Barral JM (2010) Slowing bacterial translation speed enhances eukaryotic protein folding efficiency. J Mol Biol 396: 1310–1318. - PubMed
-
- Yanofsky C (1981) Attenuation in the control of expression of bacterial operons. Nature 289: 751–758. - PubMed
-
- Chartrand P, Meng XH, Huttelmaier S, Donato D, Singer RH (2002) Asymmetric sorting of ash1p in yeast results from inhibition of translation by localization elements in the mRNA. Mol Cell 10: 1319–1330. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
