

Calcium induced regulation of skeletal troponin--computational insights from molecular dynamics simulations
- PMID: 23554884
- PMCID: PMC3598806
- DOI: 10.1371/journal.pone.0058313
Calcium induced regulation of skeletal troponin--computational insights from molecular dynamics simulations
Abstract
The interaction between calcium and the regulatory site(s) of striated muscle regulatory protein troponin switches on and off muscle contraction. In skeletal troponin binding of calcium to sites I and II of the TnC subunit results in a set of structural changes in the troponin complex, displaces tropomyosin along the actin filament and allows myosin-actin interaction to produce mechanical force. In this study, we used molecular dynamics simulations to characterize the calcium dependent dynamics of the fast skeletal troponin molecule and its TnC subunit in the calcium saturated and depleted states. We focused on the N-lobe and on describing the atomic level events that take place subsequent to removal of the calcium ion from the regulatory sites I and II. A main structural event - a closure of the A/B helix hydrophobic pocket results from the integrated effect of the following conformational changes: the breakage of H-bond interactions between the backbone nitrogen atoms of the residues at positions 2, 9 and sidechain oxygen atoms of the residue at position 12 (N(2)-OE(12)/N(9)-OE(12)) in sites I and II; expansion of sites I and II and increased site II N-terminal end-segment flexibility; strengthening of the β-sheet scaffold; and the subsequent re-packing of the N-lobe hydrophobic residues. Additionally, the calcium release allows the N-lobe to rotate relative to the rest of the Tn molecule. Based on the findings presented herein we propose a novel model of skeletal thin filament regulation.
Conflict of interest statement
Figures




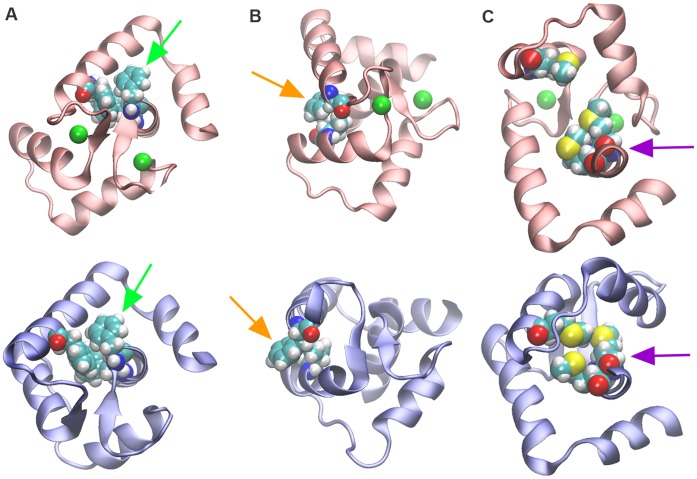
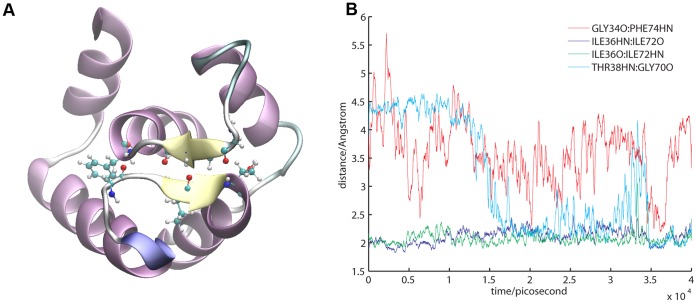
References
-
- Kobayashi T, Solaro RJ (2005) Calcium, thin filaments, and the integrative biology of cardiac contractility. Annu Rev Physiol 67: 39–67. - PubMed
-
- Arteaga GM, Kobayashi T, Solaro RJ (2002) Molecular actions of drugs that sensitize cardiac myofilaments to Ca2+. Ann Med 34: 248–258. - PubMed
-
- Endoh M (2008) Cardiac Ca2+ signaling and Ca2+ sensitizers. Circ J 72: 1915–1925. - PubMed
-
- McCammon JA, Gelin BR, Karplus M (1977) Dynamics of folded proteins. Nature 267: 585–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
