

Construction of direction selectivity through local energy computations in primary visual cortex
- PMID: 23554913
- PMCID: PMC3598900
- DOI: 10.1371/journal.pone.0058666
Construction of direction selectivity through local energy computations in primary visual cortex
Abstract
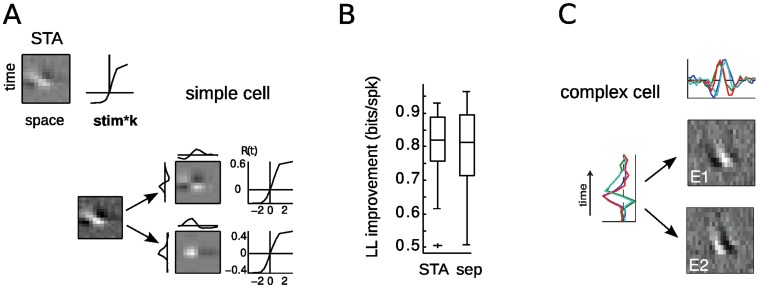
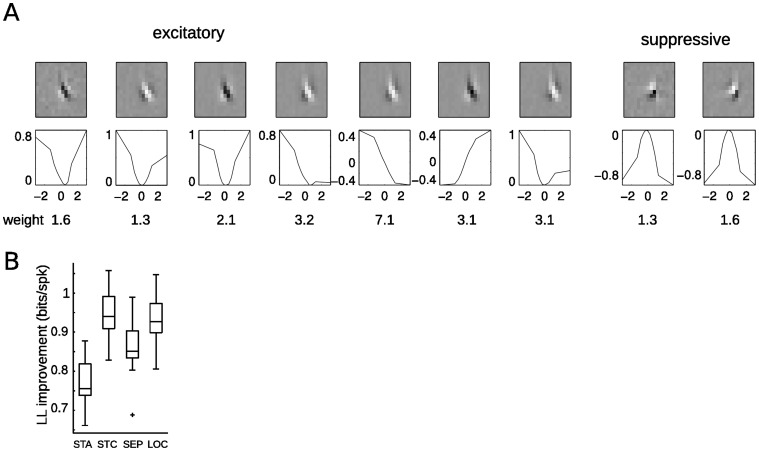
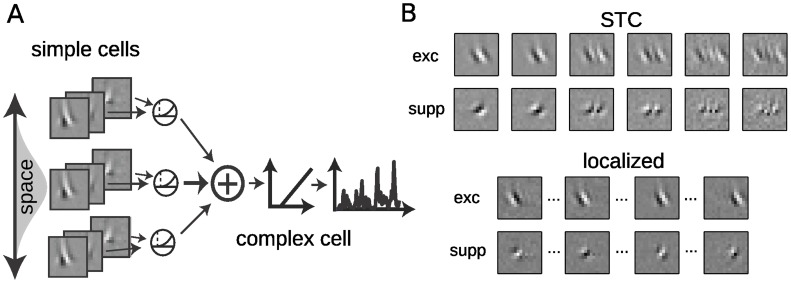
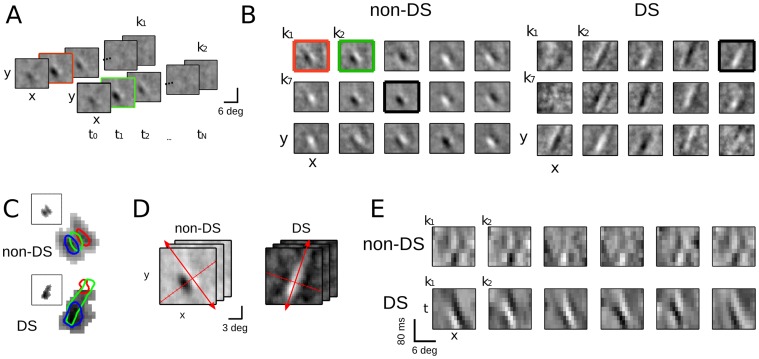
Despite detailed knowledge about the anatomy and physiology of neurons in primary visual cortex (V1), the large numbers of inputs onto a given V1 neuron make it difficult to relate them to the neuron's functional properties. For example, models of direction selectivity (DS), such as the Energy Model, can successfully describe the computation of phase-invariant DS at a conceptual level, while leaving it unclear how such computations are implemented by cortical circuits. Here, we use statistical modeling to derive a description of DS computation for both simple and complex cells, based on physiologically plausible operations on their inputs. We present a new method that infers the selectivity of a neuron's inputs using extracellular recordings in macaque in the context of random bar stimuli and natural movies in cat. Our results suggest that DS is initially constructed in V1 simple cells through summation and thresholding of non-DS inputs with appropriate spatiotemporal relationships. However, this de novo construction of DS is rare, and a majority of DS simple cells, and all complex cells, appear to receive both excitatory and suppressive inputs that are already DS. For complex cells, these numerous DS inputs typically span a fraction of their overall receptive fields and have similar spatiotemporal tuning but different phase and spatial positions, suggesting an elaboration to the Energy Model that incorporates spatially localized computation. Furthermore, we demonstrate how these computations might be constructed from biologically realizable components, and describe a statistical model consistent with the feed-forward framework suggested by Hubel and Wiesel.
Conflict of interest statement
Figures
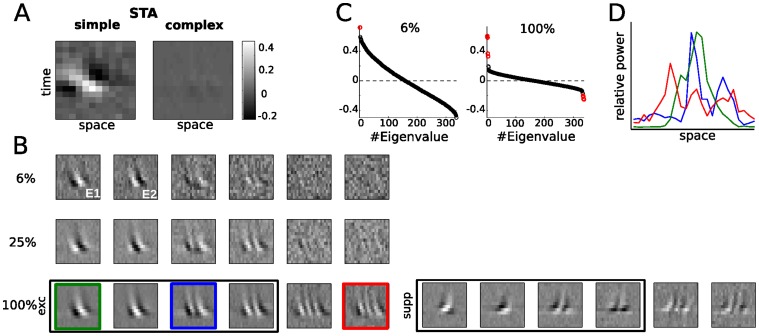
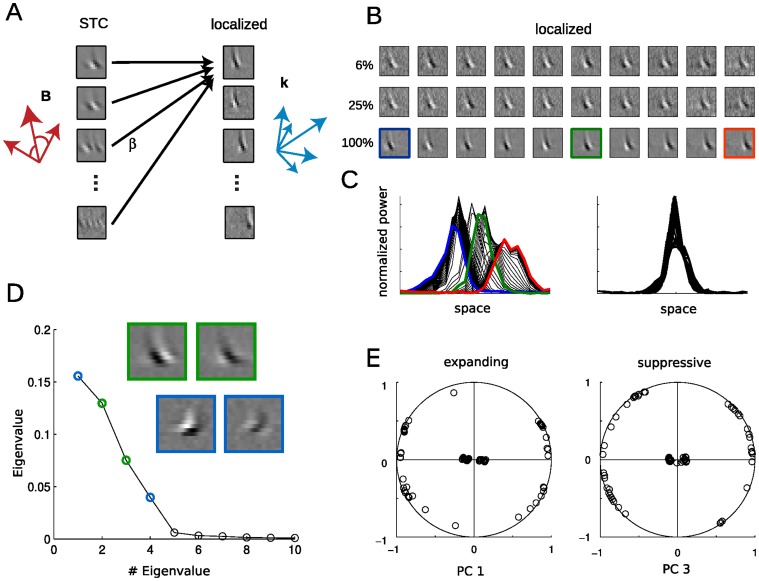
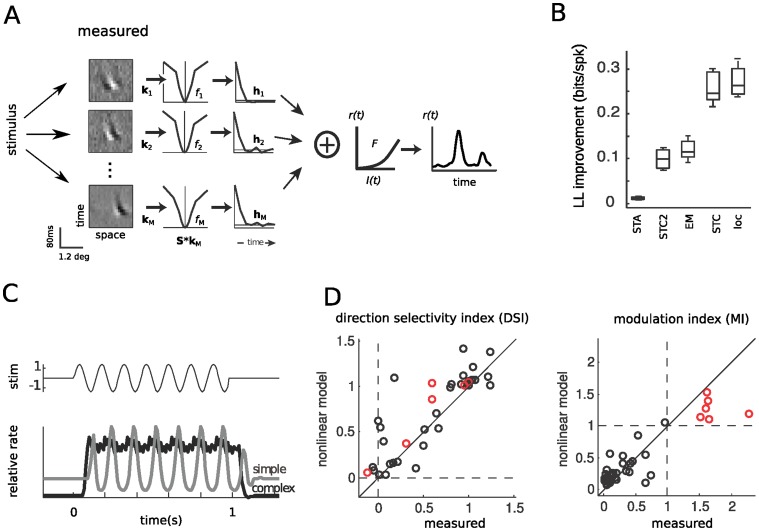
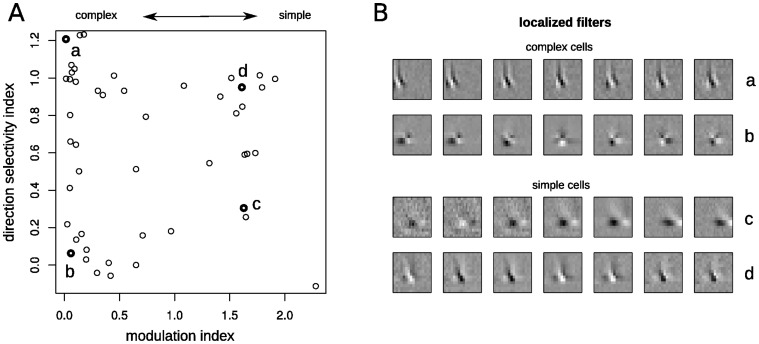
Similar articles
-
Model-based characterization of the selectivity of neurons in primary visual cortex.J Neurophysiol. 2022 Aug 1;128(2):350-363. doi: 10.1152/jn.00416.2021. Epub 2022 Jun 29. J Neurophysiol. 2022. PMID: 35766377 Free PMC article.
-
A Computational Model of Direction Selectivity in Macaque V1 Cortex Based on Dynamic Differences between On and Off Pathways.J Neurosci. 2022 Apr 20;42(16):3365-3380. doi: 10.1523/JNEUROSCI.2145-21.2022. Epub 2022 Mar 3. J Neurosci. 2022. PMID: 35241489 Free PMC article.
-
Direction selectivity of synaptic potentials in simple cells of the cat visual cortex.J Neurophysiol. 1997 Nov;78(5):2772-89. doi: 10.1152/jn.1997.78.5.2772. J Neurophysiol. 1997. PMID: 9356425
-
Surround suppression supports second-order feature encoding by macaque V1 and V2 neurons.Vision Res. 2014 Nov;104:24-35. doi: 10.1016/j.visres.2014.10.004. Epub 2014 Oct 23. Vision Res. 2014. PMID: 25449336 Free PMC article. Review.
-
Mechanisms of neuronal computation in mammalian visual cortex.Neuron. 2012 Jul 26;75(2):194-208. doi: 10.1016/j.neuron.2012.06.011. Neuron. 2012. PMID: 22841306 Free PMC article. Review.
Cited by
-
Spike-Triggered Covariance Analysis Reveals Phenomenological Diversity of Contrast Adaptation in the Retina.PLoS Comput Biol. 2015 Jul 31;11(7):e1004425. doi: 10.1371/journal.pcbi.1004425. eCollection 2015 Jul. PLoS Comput Biol. 2015. PMID: 26230927 Free PMC article.
-
Mapping nonlinear receptive field structure in primate retina at single cone resolution.Elife. 2015 Oct 30;4:e05241. doi: 10.7554/eLife.05241. Elife. 2015. PMID: 26517879 Free PMC article.
-
Temporal Limits of Visual Motion Processing: Psychophysics and Neurophysiology.Vision (Basel). 2019 Jan 26;3(1):5. doi: 10.3390/vision3010005. Vision (Basel). 2019. PMID: 31735806 Free PMC article.
-
Cortical direction selectivity increases from the input to the output layers of visual cortex.PLoS Biol. 2025 Jan 8;23(1):e3002947. doi: 10.1371/journal.pbio.3002947. eCollection 2025 Jan. PLoS Biol. 2025. PMID: 39777916 Free PMC article.
-
Model-based characterization of the selectivity of neurons in primary visual cortex.J Neurophysiol. 2022 Aug 1;128(2):350-363. doi: 10.1152/jn.00416.2021. Epub 2022 Jun 29. J Neurophysiol. 2022. PMID: 35766377 Free PMC article.
References
-
- Van Essen DC, Anderson CH, Felleman DJ (1992) Information processing in the primate visual system: an integrated systems perspective. Science 255: 419–423. - PubMed
-
- Jia H, Rochefort NL, Chen X, Konnerth A (2010) Dendritic organization of sensory input to cortical neurons in vivo. Nature 464: 1307–1312. - PubMed
-
- Rust NC, Stocker AA (2010) Ambiguity and invariance: two fundamental challenges for visual processing. Curr Opin Neurobiol 20: 382–388. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
